The beetles and the beasts

Material & methods
Study Area
The study area is the Nordhordland Biosphere Reserve (NBR), including nine grassland study sites (Figure 1). Three of the sites were sampled closer to the coast (O’ coding/outer fjord, Table 1), while the other six sites were in the fjords (I’ coding/coast, Table 1). All sites were grazed by either cows, sheep, or goats. Additional details are listed in Table 1. We collected the samples in systems with low-intensity grazing, and low to no pesticide usage.
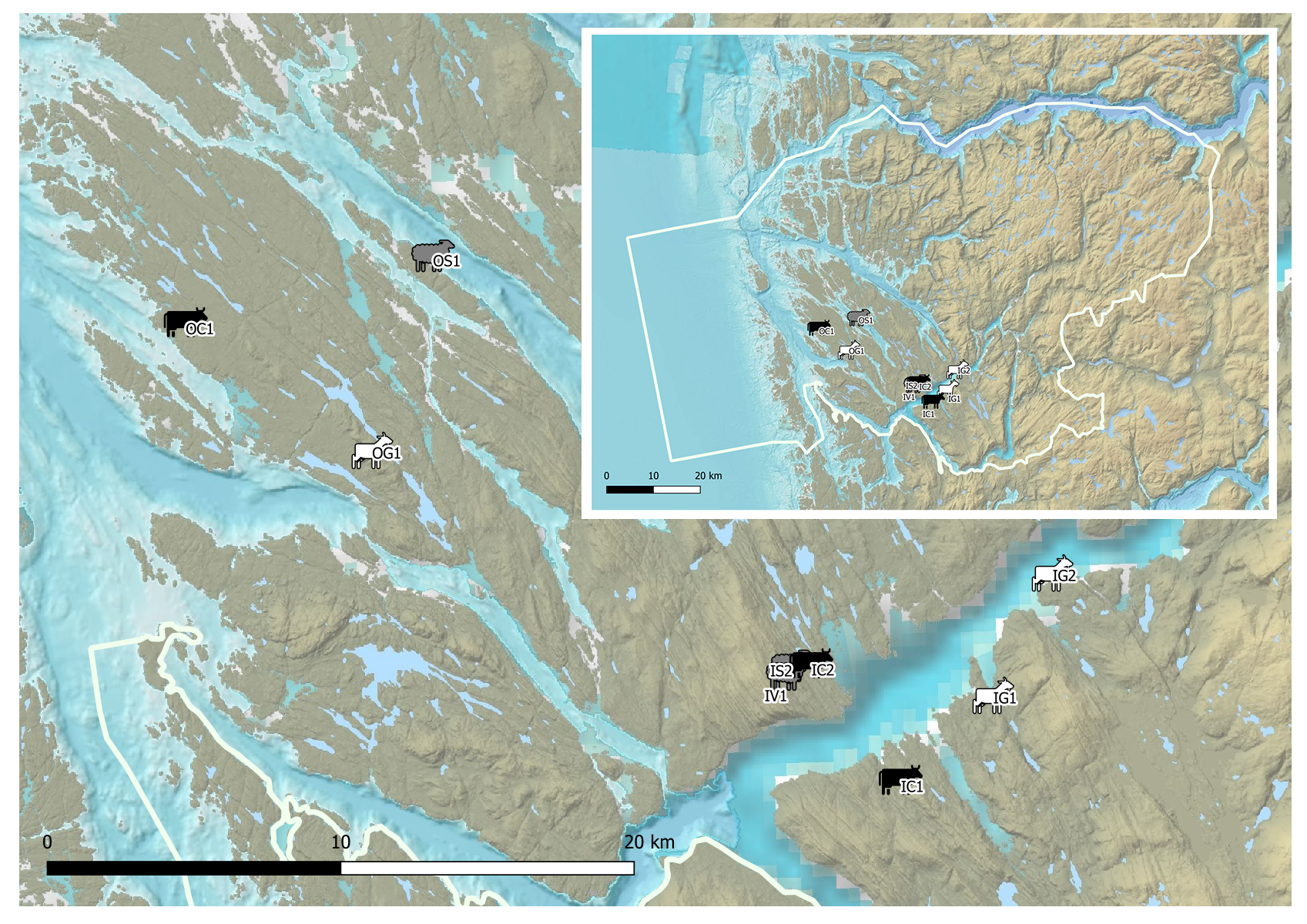
Figure 1. Location of the nine sites grazed either by sheep (grey), goat (white) or cows (black). Credits: Morgane Kerdoncuff.
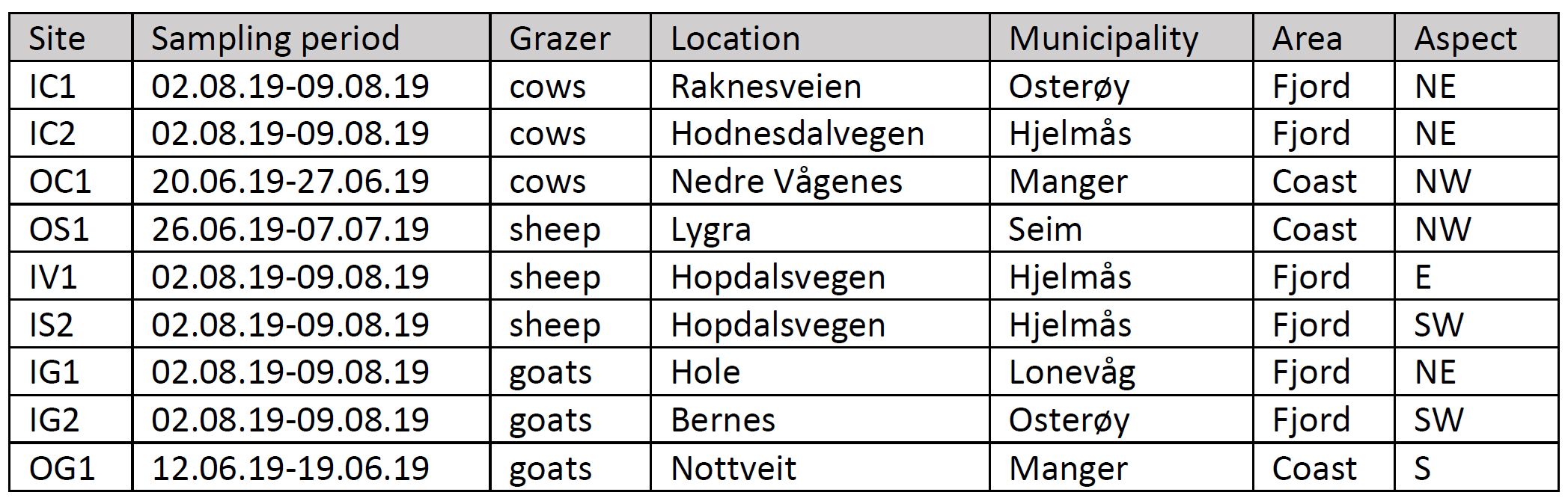
Table 1. Description of the nine study sites in Nordhordland Biosphere Reserve.
Data Collection
Sites were defined as sampling areas of 20 x 20 m2, with homogeneous grassland vegetation cover representative of the surrounding grazing field area. Wet zones (e.g., with peat moss or cotton grass) or bare rock were avoided. The aspect and elevation of the sites were recorded. In each site, we installed three sets of four dung-baited pitfall traps. One set was made of a 1 m2 square, with one trap on each corner. Sets were separated from each other by a minimum distance of 6 m2. There was a total of 12 traps per site, which resulted in 108 samples for the overall survey. Each trap had a diameter of 11.5 cm and was covered by a wire mesh and baited with cow dung to attract dung beetles.
The samples in this project are from different periods during summer 2019, with traps being active for one week each (Table 1). All beetles were then sorted by family in the lab and stored in 70 % ethanol. For family identification I used Duff & Smith (2012) and Unwin (1985).
Data Handling
The survey will address the following response variables: total abundance of beetles, abundance per family and family diversity. The variables were visualized as bar plots using Microsoft Excel.
Results
My results showed that different grazers had different impacts on beetle communities. It seems that beetle abundance may not be related to beetle diversity.
The abundance of beetles per. site ranged from 69 (IG2) to 1672 (OS1) individuals (Figure 2). The sites grazed by sheep had an overall greater abundance of beetles but also showed the highest variability across sites. The lowest abundance of beetles is found in a goat site (IG2). On average I found a total abundance of beetles of 157 for sheep, 97 for goats, and 77 for cows. There is a clear difference between fjord and coastal sites, especially for sheep and goats – reflected in e.g., four times as many beetles in the OS1 site than in the IS2 site.
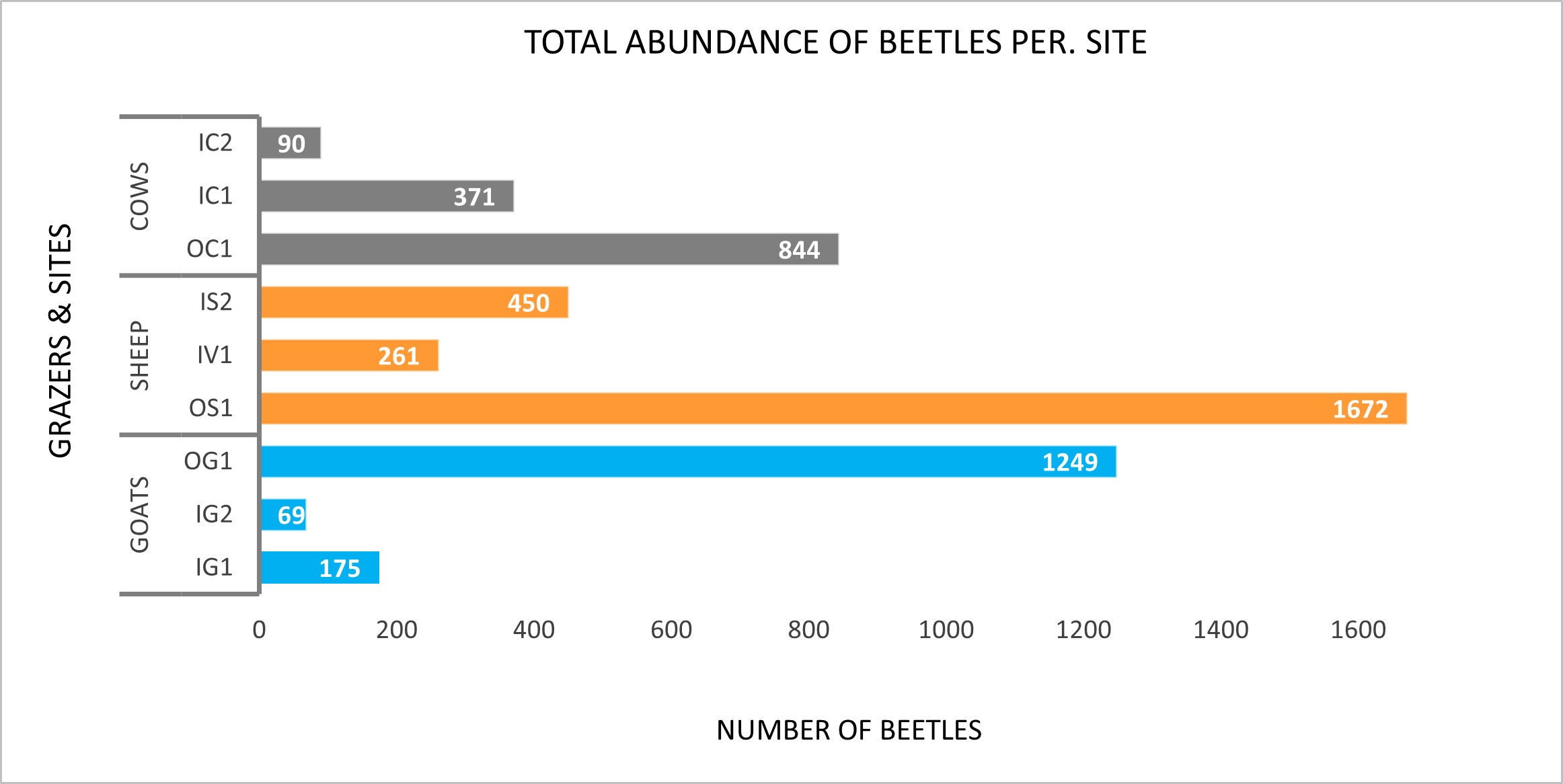
Figure 2. Total abundance of beetles for each site, classified according to livestock type (cows, sheep, goats).
The coastal sites (OC1, OS1, OG1) had an overall higher abundance of beetles, particularly for Staphylinidae (rove beetles) but also for Hydrophilidae (water scavenger beetles) (Figure 3). Staphylinids are most abundant for all sites, except the IC2 site grazed by cows. Carabidae (ground beetles) and Scarabidae (dung beetles) are the least abundant families throughout nearly all sites. The sheep sites IS2 and OS1 stood out by having the highest abundance of dung beetles. The goat sites in fjords had the lowest abundance of beetles per. family of all the fjord sites in the survey.
The total abundance of beetles is highest in sheep sites (Figure 2). These sites also have the highest abundance of beetles in different families (Figure 3). This is true for three out of the five families analysed, respectively the families of Scarabidae, Staphylinidae, and Hydrophilidae. Ptiliidae (featherwing beetles) were most abundant in OC1 (Figure 3), however, they seemed to thrive in any habitat.
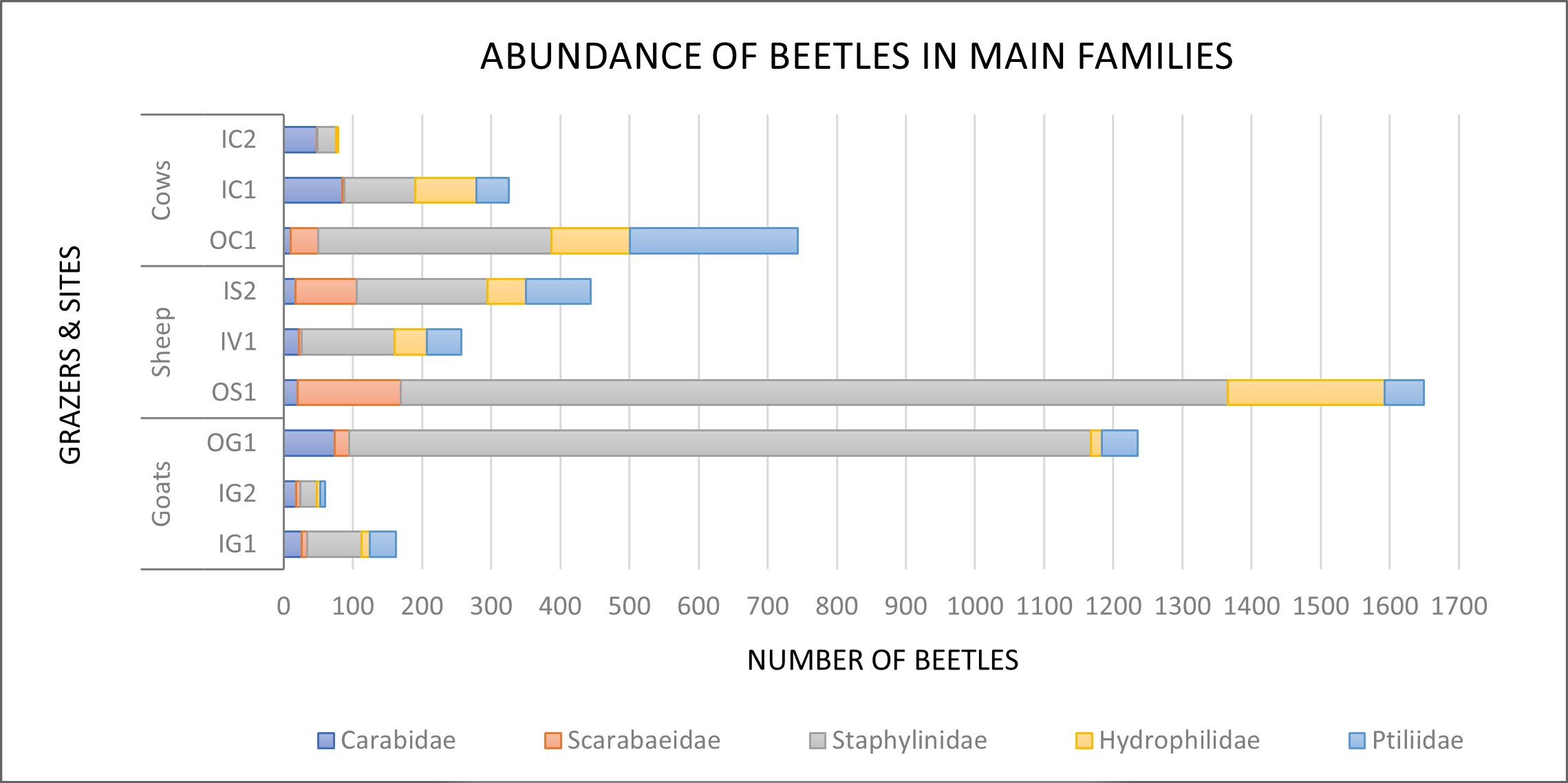
Figure 3. Total abundance of beetles for the main families (Carabidae, Scarabaeidae, Staphylinidae, Hydrophilidae and Ptiliidae) in each site, classified by livestock type.
The diversity ranges from 5.0 to 9.0 families per site. Cow and goat sites have 9.0 families in two out of three sites. The average number of families is highest in cow sites by 8.0, closely followed by goats with 7.7 and sheep sites with an average of 7.0 families per site (Figure 4).
From figure 2, 3 and 4 we can observe that beetle abundance and diversity do not always follow the same trends. For instance, beetle abundance is substantially lower on IG2 and IC1, yet the number of families identified is quite similar to other sites.
Regarding geographical location, the abundance in main families is highest in coastal sites – as with the overall abundance.
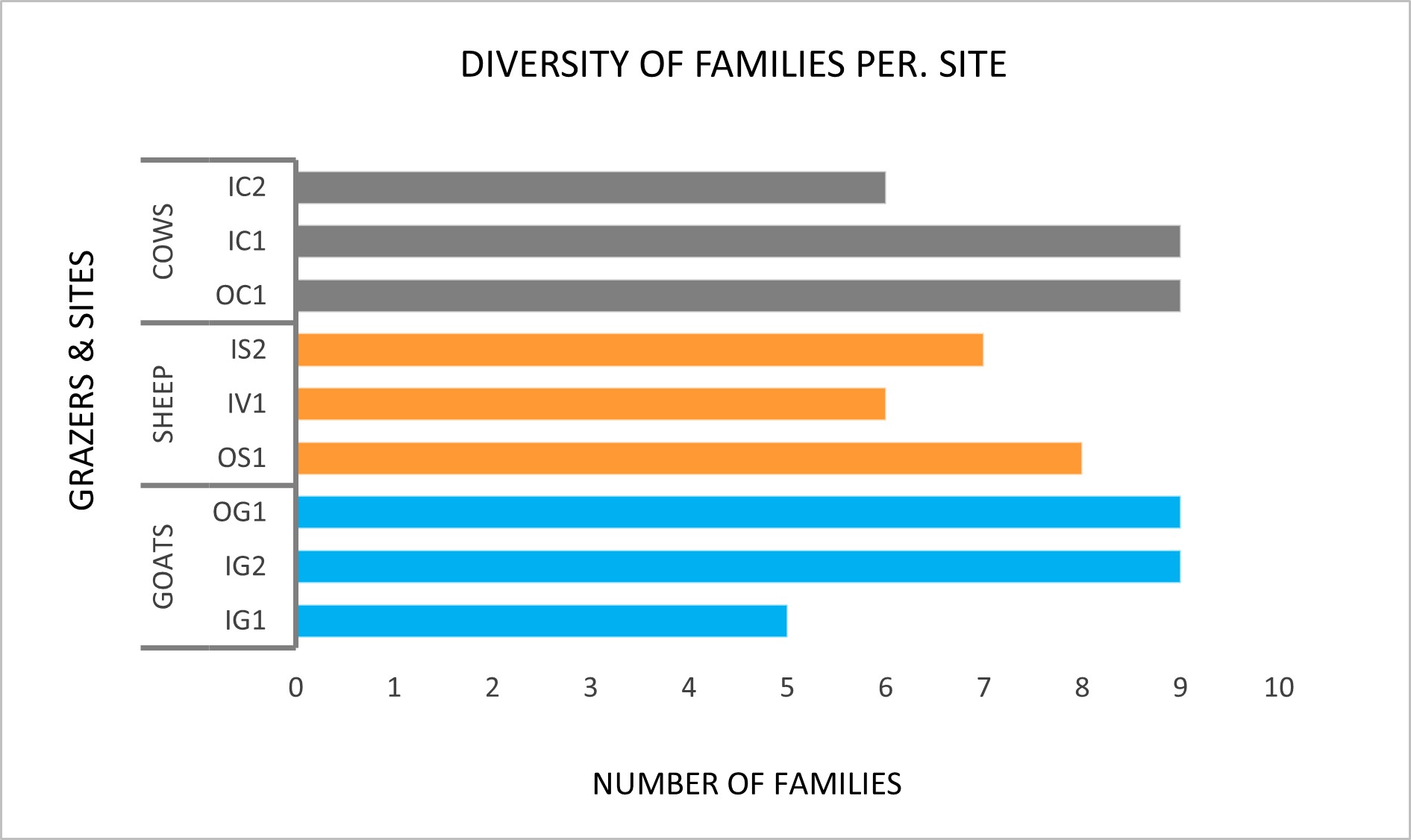
Figure 4. Total number of beetle families per site, classified according to livestock type.
About the author

Ingrid Vaksvik
Hi! I am a 38-year young lady from a small village in Sunnmøre, Western Norway. My greatest passion in life are insects, and they have fascinated me for as long as I can remember. It was therefore a natural choice to take a master's degree in biology, with a priority on insects. In addition, I am also interested in plants, sustainability, nature management and agriculture – something I have learned more about during my studies, but which I also got to use in my contribution to Bikuben. My text is a revised article that was prepared in connection with the topic “BIO299 - Research Practice.” The experience from the course as well as the writing process with through Bikuben has given me a taste for academic writing, laboratory work and fieldwork. This type of holistic work methodology is something I want to combine with a future job in terrestrial biology. My aim with the study and not least with this article is to pay more attention to the bizarre but important insects that surround us wherever we go, but which we often take for granted. They are perhaps best known for giving us beautiful flowers, honey, strawberries, apple, almonds and many other species of berries, fruits, and vegetables. But insects are more than just pollinators; They are nature's own waste collectors and can be indicators of the health of an ecosystem. In addition, they are important for birds and fish - something that humans also benefit from. Not least, the insects entertain us with a mysterious world full of crazy and ingenious inventions.
The structures between the biotic and the abiotic; air, light, water, soil, plants, insects, and other animals are complex and vulnerable at the same time. If one step on the ladder of life changes, it can have fatal consequences for subsequent steps, and so on. The world will in many ways be a poorer place without the insects. We need increased knowledge and commitment to protect these creatures – both among decision-makers and the general population. My article is an attempt to raise this awareness and I hope my contribution might make a difference.

Credits: Ingrid Vaksvik



1 Response
[…] I. (2023). The beetles and the beasts. Bikuben […]