Characterizing Expression Patterns of LexA and LexAop Lines for the Development of a Drosophila Cancer Model

Materials & methods
At the University of Bergen's fly lab facility, flies were cultivated in an incubator at 25 °C and with a light regulation that mimicked the daily rhythm of the sun in the flies' natural environment. The light turns on at 9.00 in the morning and off at 9.00 in the evening. Flies were stored in tubes containing a food medium and were flipped1 once a week to a fresh tube of food. All different genetic fly lines were provided by Fergal O’Farrell, associate professor at the University of Bergen (Table 1). Table 1 abbreviations will be used to refer to the genetic lines throughout this paper.
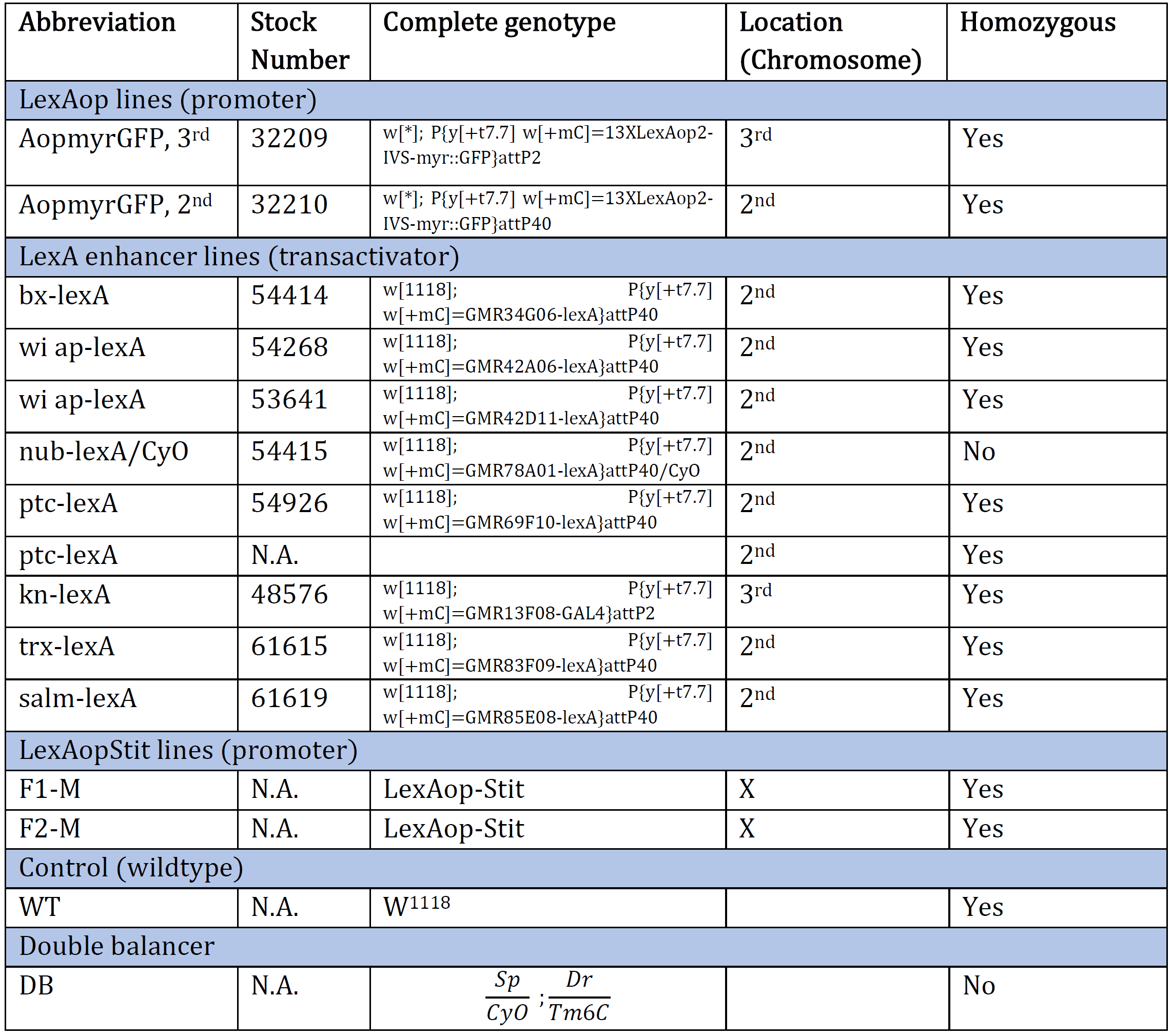
Table 1: An overview of the Drosophila genotype of original genetic lines provided at the beginning of the experiment. The genotype and the corresponding stock number are aligned. The table also provides an abbreviation for each genotype and which chromosome the gene of interest is inserted/present. N.A. indicates information Not Available.
Selection of Flies - Fly Pushing
The balancers had different phenotypic expressions, enabling us to sort out the flies containing the genetic lines of interest with the use of a microscope (Figure 1). The technique used is called fly pushing and refers to the daily sorting of flies. Flies are emptied from their tube onto a CO2-perfused pad, knocking out the flies instantly. A small paintbrush is then used to ‘push’ the flies around to select for or against anatomical characteristics and phenotypic expressions. A Leica light microscope with additional spotlights was used for fly pushing.
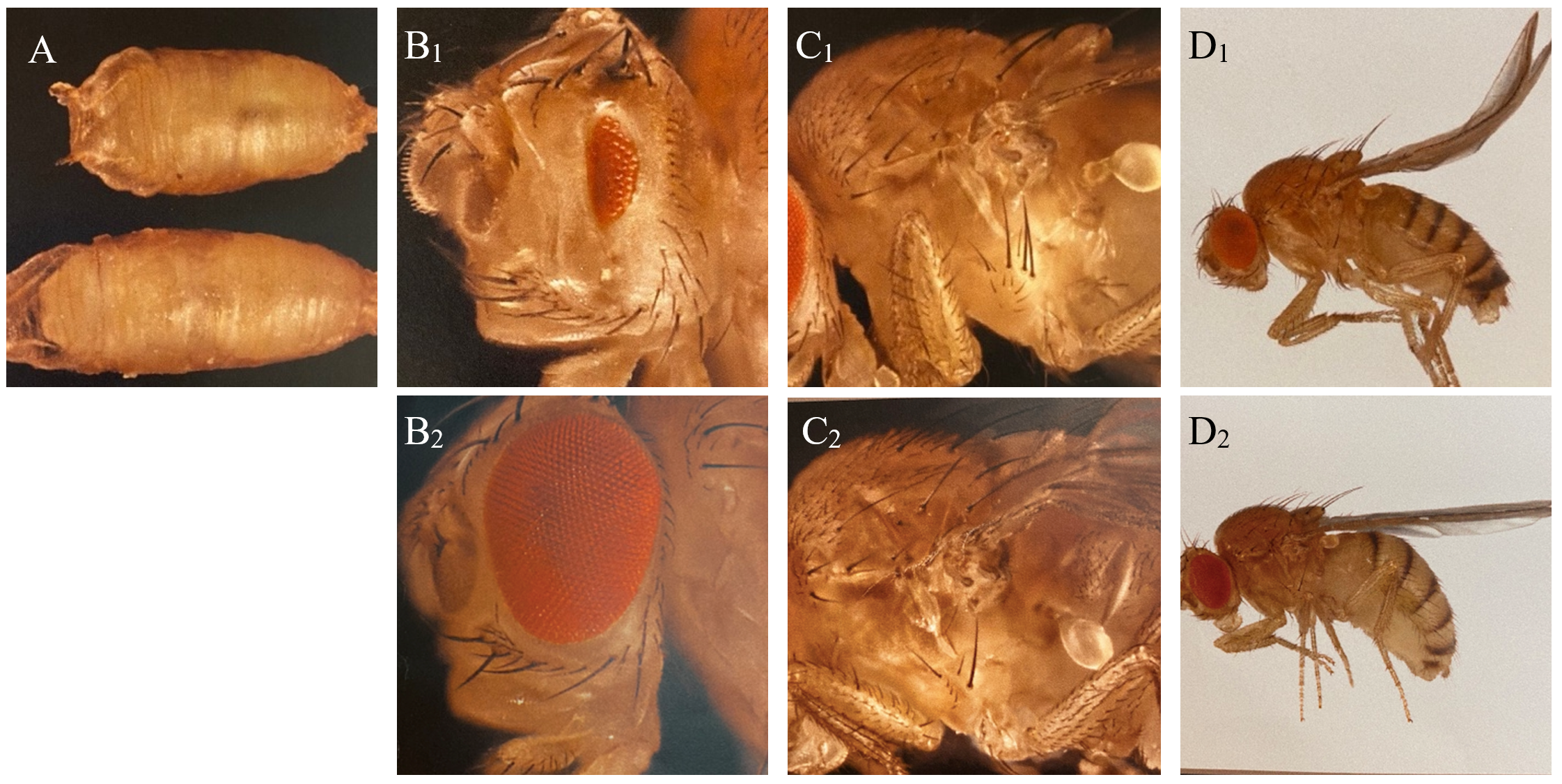
Figure 1: Phenotypic expressions of balancers and corresponding wild type of Drosophila used in the experiment. A: C: Tb (Tm6C), reduction of body size (above), and wildtype (below). B: Dr, reduction of eye (B1) and wildtype (B2). C: Sp, additional hairs behind the first anterior leg of the fly (C1) and wildtype (C2). D: CyO, curly wings (D1) and wildtype (D2).
Setting Crosses
Crosses were set on a Friday, ensuring that the F1 generation started to emerge on a Monday (10 days after fertilization). In order to make a cross, 4-8 virgin females and 2-4 males were required. Virgin females have specific characterisations visible with the use of a microscope. They are more transparent, paler in colour, and possess bloated abdomens. The crosses were flipped to a new tube 4-5 days after the cross had been made, preventing the parent generation and F1 generation to be mixed.
Generation of Double-Balanced Stocks
The goal was to create double-balanced stocks containing a LexA enhancer line on the 2nd chromosome, and LexAopmyr::GFP 3rd chromosome (Figure 2). There was one exception, that of the KnLexA, where the aim was to have both the LexA and LexAopmyr::GFP on the 3rd chromosome. The differentiation was caused due to the position of the KnLexA enhancer on the 3rd chromosome (Figure 2). The goals required a multistep crossing scheme. A short summary of the original genetic lines needed to create each double-balanced stock is included (Figure 2). A slash (/) indicates that the genes are on the same chromosome whereas a semicolon (;) indicates that the genes are on separate chromosomes of Drosophila.
Simultaneously as the multistep process of creating double-balanced stocks started, direct crosses of each LexA promoter line and Aopmyr::GFP were also made (before the enhancers had been balanced). This was done to check if there was a GAL4 expression pattern, shown with GFP-positive cells if present. If the results were negative, meaning no expression pattern, the given LexA enhancer line would not be used further in the experiment.
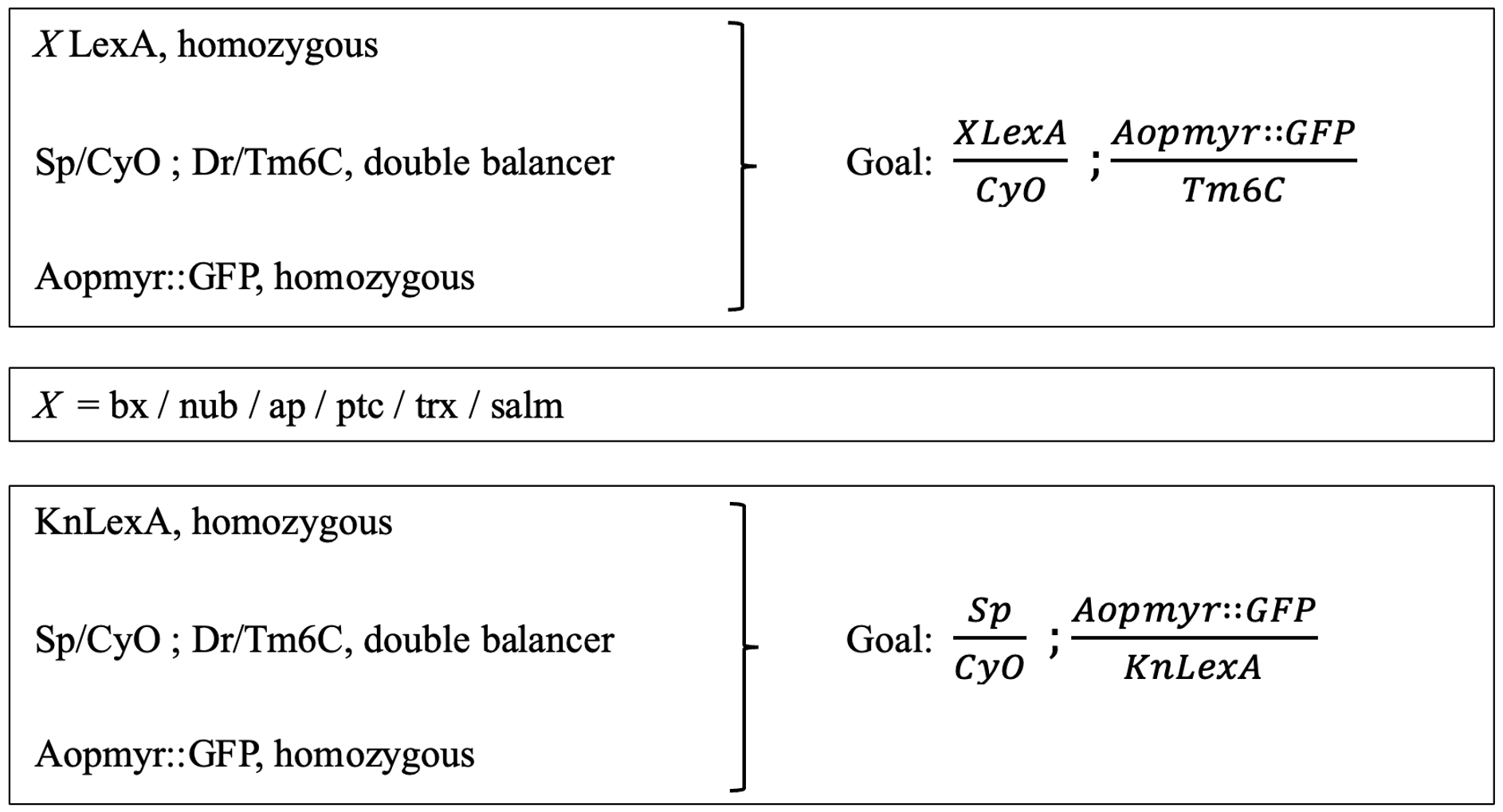
Figure 2: Presentation of the crossing scheme aim for obtaining double balanced stocks. The original lines (left) that were used to create the goal (right). The procedure was consistent for all different LexA enhancer lines; therefore, the X is used in order to indicate each LexA enhancer line. The goal and original lines needed for KnLexA are also shown.
Microscopy and Dissection
The imaginal wing disk of wandering third-instar larvae was removed by dissection and fixed in 500 µl 4% formaldehyde/PBS (Phosphate-Buffered Saline) for 20 minutes in a well plate. Then the disks were rinsed with PBS for 3x1 minute before being fixed in 500µl PBS containing Hoechst 10 µg/ml with a 1:10 000 ratio for 20 minutes. Lastly, the disks were rinsed with PBS.
A drop of glycerol mountant was added to an objective slide by using a pipette. The imaginal wing disks were then transferred to the mountant, and an objective glass was placed on top. The objective slides were labelled immediately afterward with the genotype of the imaginal disks. All objectives were stored in a fridge until microscopy of the disks. The dissection was performed using a surgical needle and forceps. A minimum of three discs showing the same result for each genotypic expression was required before conclusions could be drawn.
Illumination microscopy imaging was performed using a Leica inverted microscope and the LasX Leica software. The disks were illuminated with 420nm and 488nm wavelength light produced by an LED laser light source for the detection of Hoechst and GFP-positive cells/expression, respectively. Following image acquisition samples were compared using ImageJ Fiji.
Double-Balanced Enhancer LexA lines exposed to Stit
The completed double-balanced LexA;AopmyrGFP stocks were crossed with LexAopStit (Stit). The goal was to detect any changes that might occur within the GAL4 expression pattern when being exposed to Stit. At least two samples of each Stit;LexA;AopmyrGFP as well as a control, not crossed to Stit, were made for each completed line following the same procedure as the microscopy and dissection before (section "Microscopy and Dissection"). Due to limited time, only the LexA enhancer lines PtcLexA and ApLexA was exposed to Stit.
Results
Direct Crosses of LexA Promoter lines for GAL4 Expression
The expression patterns of all results are shown with GFP-positive cells, if present, within the imaginal wing disc of Drosophila. The direct cross of different LexA promoter lines for GAL4 expression is presented in Figures 3 – 5. GFP-positive cells were observable in the LexA promoter fragmented genetic lines of Trx, Ap, Ptc, and Nub (Figure 4 – 5), whereas the LexA promoter fragmented genetic lines Bx, Kn, and Salm were GFP-negative (Figure 3). The GFP-positive expression sites varied within the imaginal wing disc among the different genetic lines, however, all lines had some expression within the wing pouch area. Further NubLexA only showed expression in 50% of the imaginal discs dissected due to this line not being homozygous. To confirm GFP-positive cells a tube with larvae was held under a LED light where GFP was visible as a brighter region in the anterior region of the larvae (Figure 6).
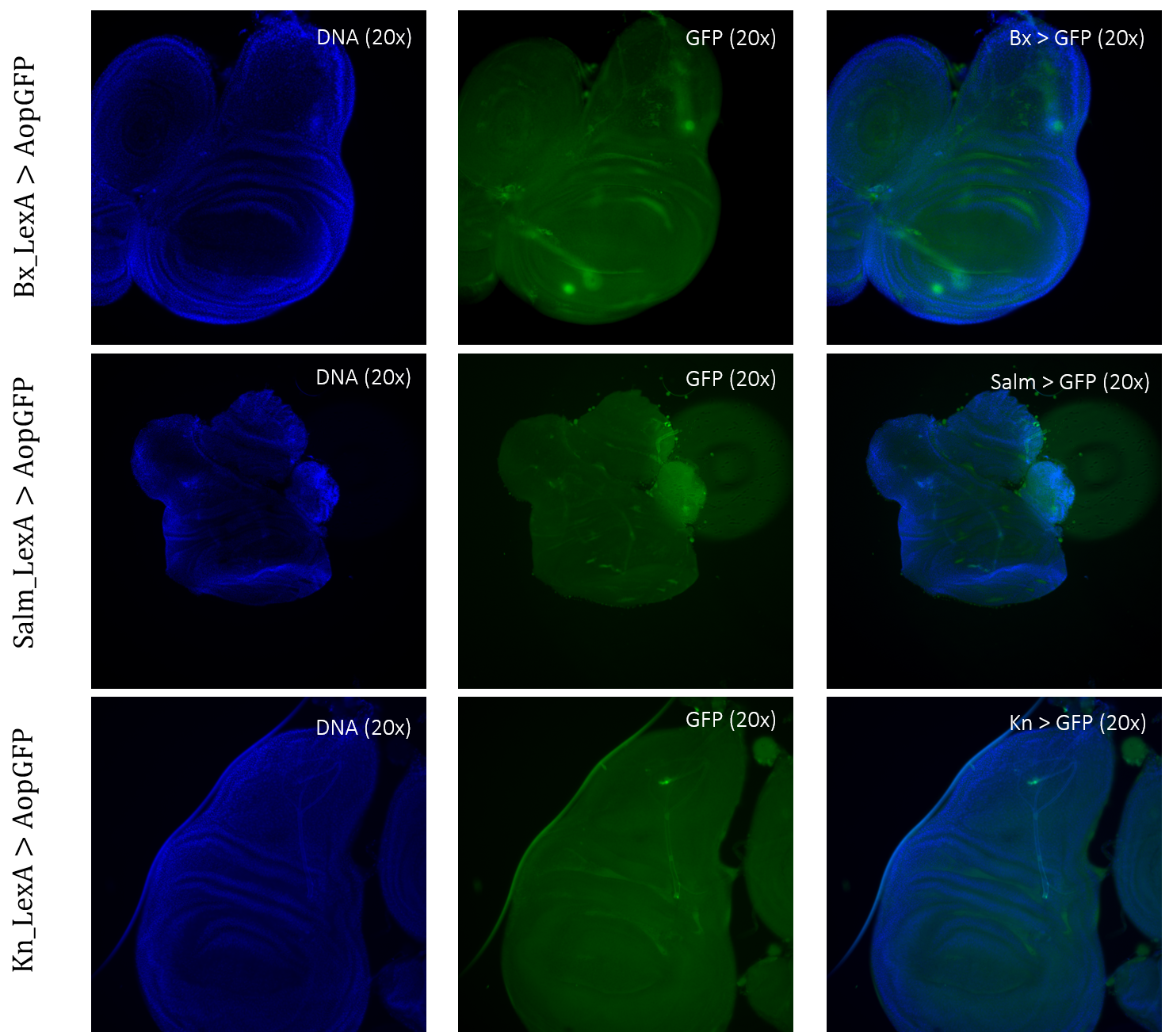
Figure 3: Negative GFP-cells in tissue samples from LexA promoter for GAL4 expression in Drosophila. BxLexA, SalmLexA, KnLexA, showed no expression sites of GFP-positive cells within the imaginal wing disk of Drosophila. The GFP scan shows that there is only background colouring for these driver lines.
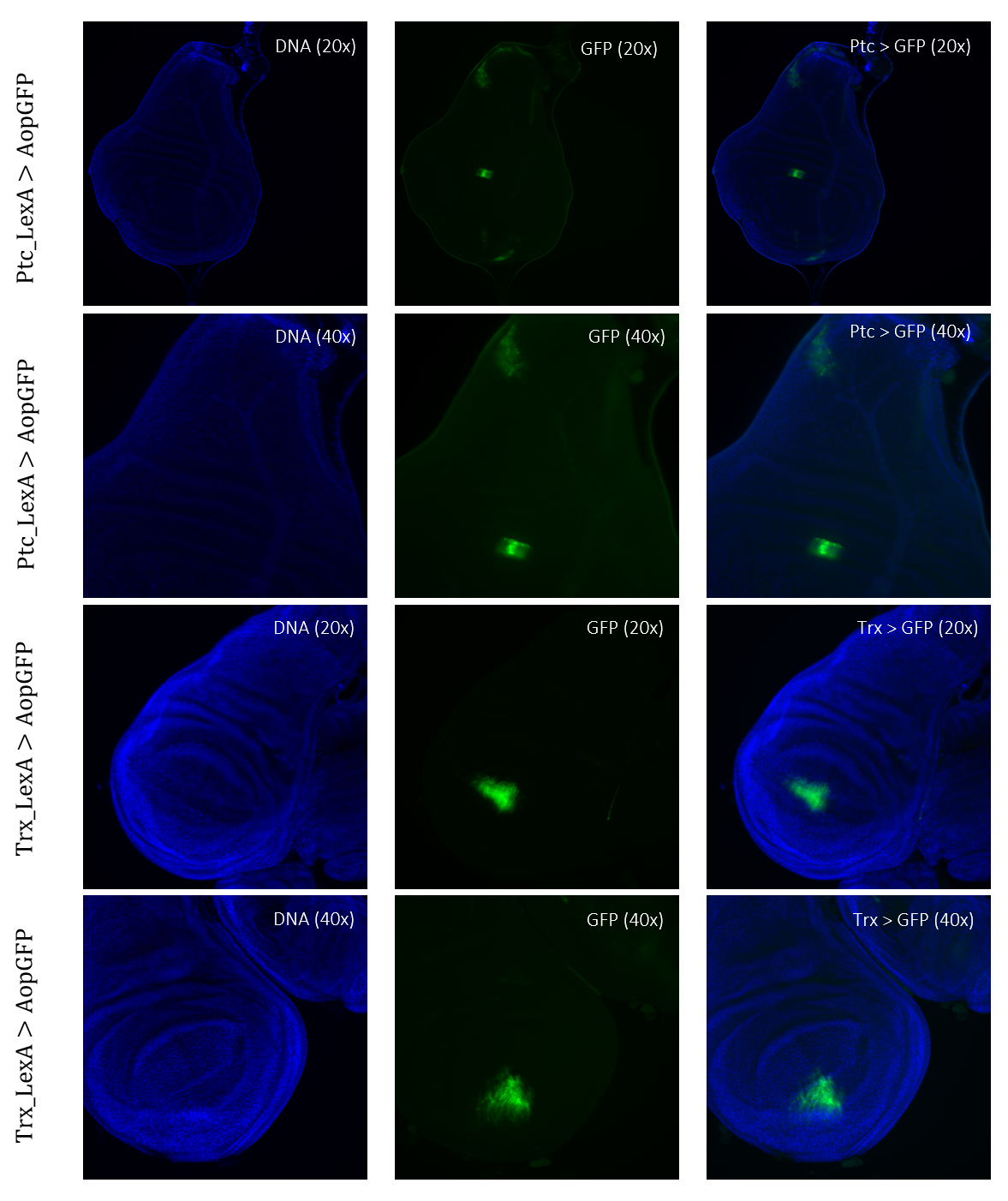
Figure 4: Tissue samples from LexA promoter for GAL4 expression in Drosophila. GFP-positive cells were observable in different regions of the imaginal wing disk of Drosophila for driver lines Ptc(54926)LexA and TrxLexA. PtcLexA showed fragmented expression within the wing disk, including the wing pouch and notch. TrxLexA showed a restricted region of positive GFP cells in the pouch area of the imaginal wing disk.
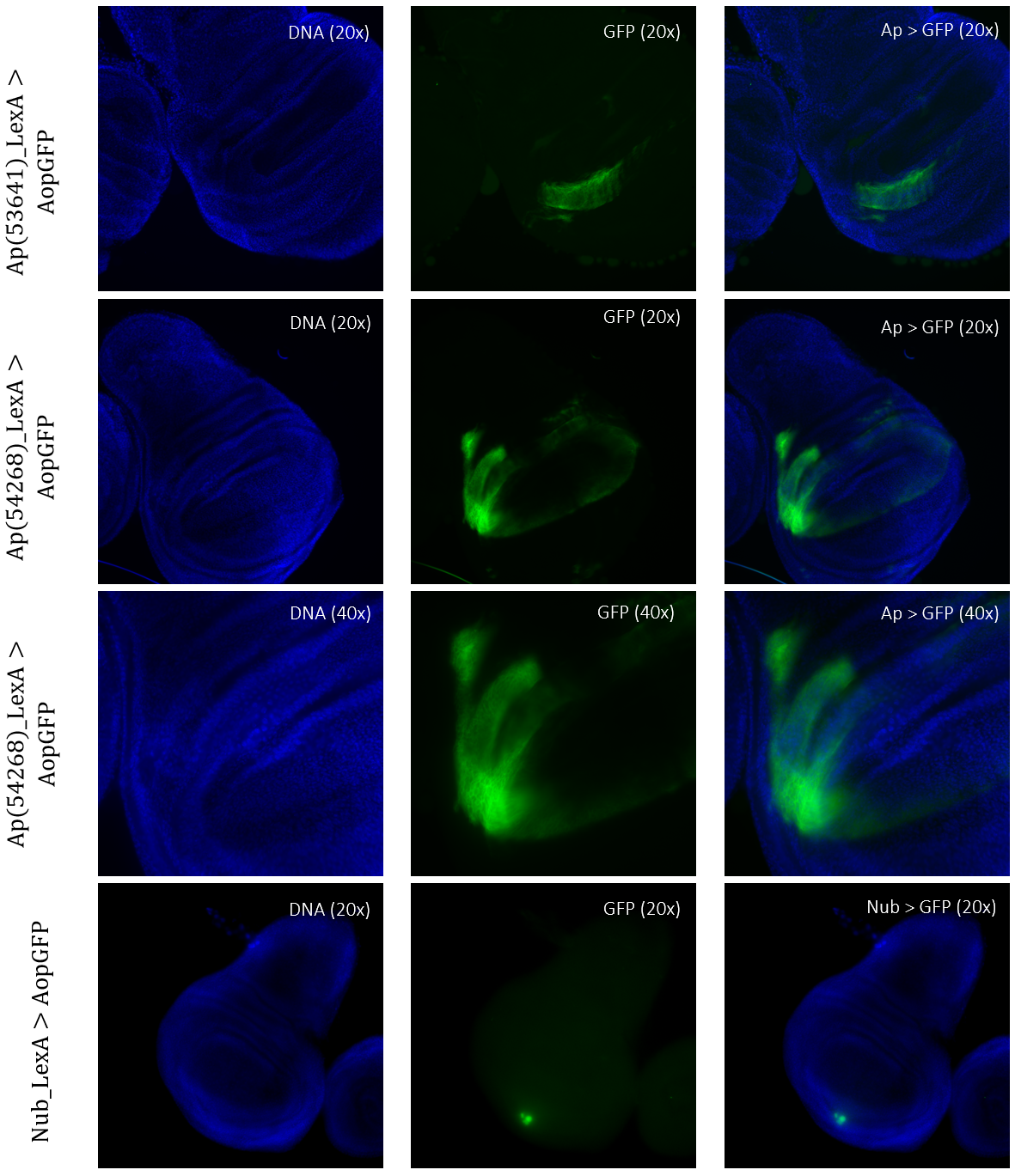
Figure 5: Tissue samples from LexA promoter for GAL4 expression in Drosophila. GFP-positive cells were observable in different regions of the imaginal wing disk of Drosophila for driver lines Ap(53641)LexA, Ap(54268)LexA, and NubLexA. Both ApLexA lines had GFP-positive cells in the dorsal region of the wing pouch, where the expression pattern of Ap(54268) was also present in the wing thorax. NubLexA showed three small fragments of GFP-positive cells within the ventral region of the wing pouch.
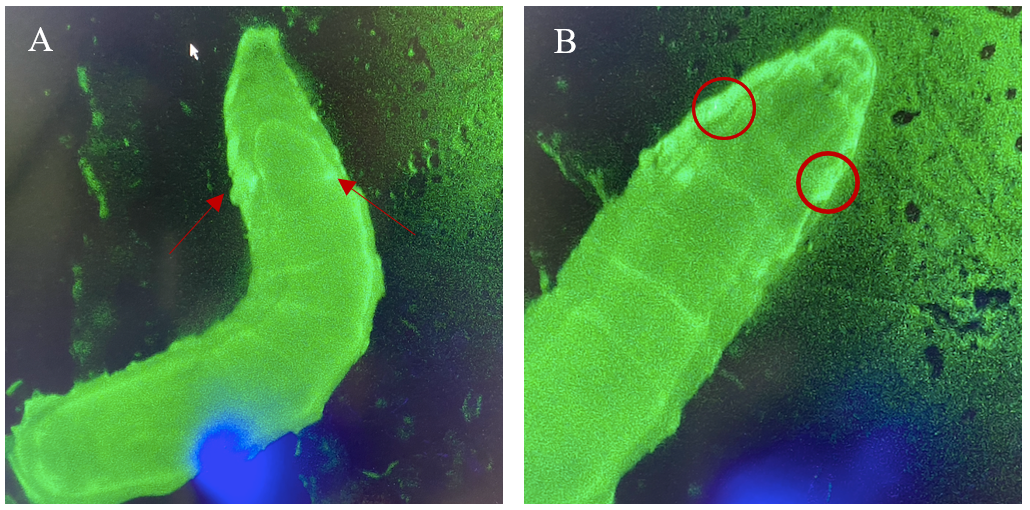
Figure 6: 3rd instar larvae with NubLexA promoter for GAL4 expression in the imaginal wing disc. The GFP-positive region was identified in the anterior region of the larvae as a small circle with a brighter colour on both the left and the right side of the organism. The figure shows one larva to the left (A) and the same larva but at a higher magnification to the right (B). The areas with expressions are highlighted with arrows and circles.
Crosses of Double-Balanced LexA promoter lines for GAL4 Expression when exposed to Stit
Crosses of double-balanced lines Ptc>GFP and Ap>GFP with Stit-transformation of cells showed abnormal expression patterns. Two different positioned Stit-transformations were used, Stit FM1 for Ptc expression (Figure 7) and Stit FM2 for Ap expression (Figure 8). Stit>Ptc>GFP, Stit-transformed cells, showed an elongated expression fragment within the wing pouch of the first sample, less centered than that of the control (Figure 7). The second sample showed more centered fragments similar to that of the control. The region of expressions for both samples were overlapping with that of the control.
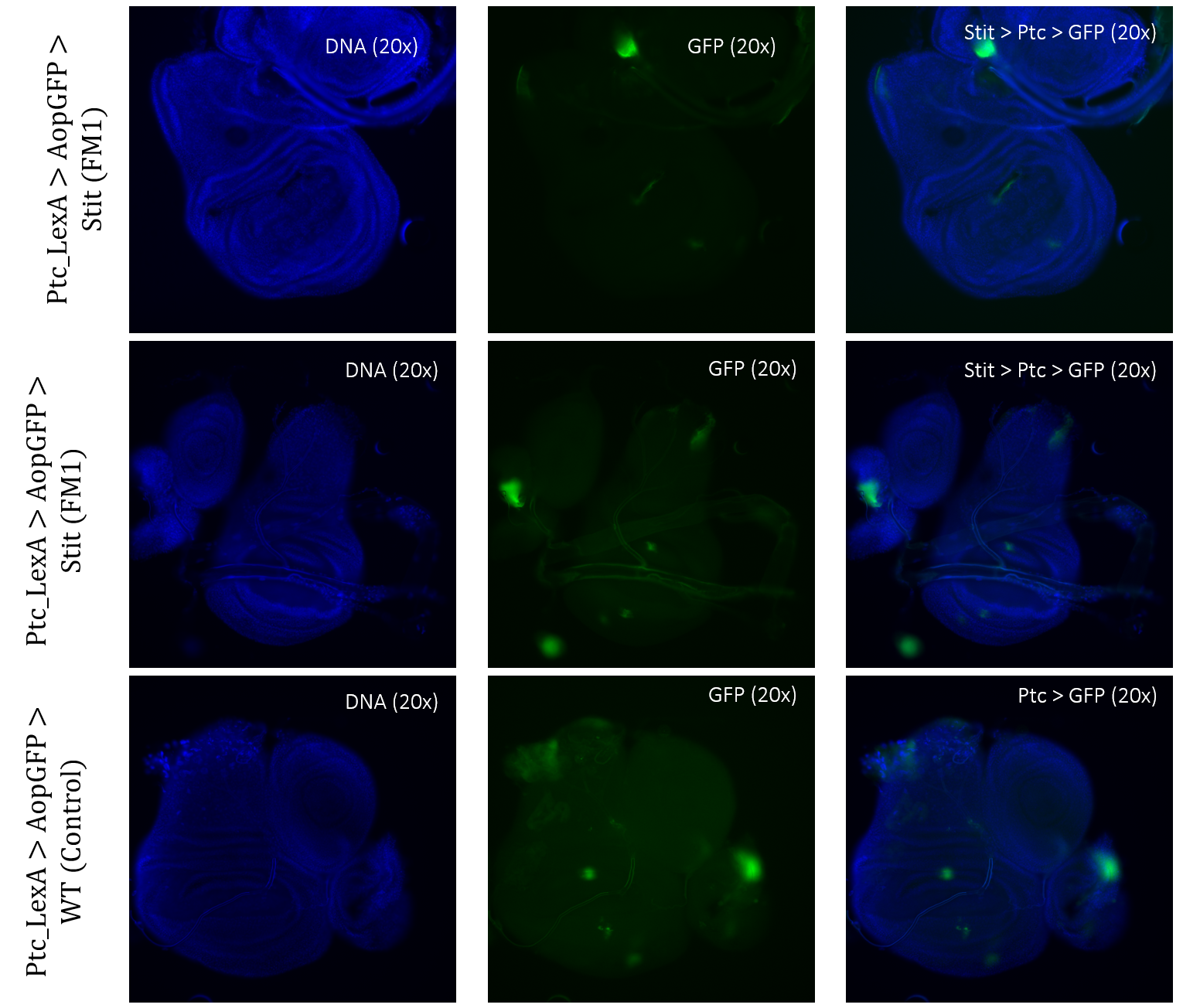
Figure 3.5: Tissue samples from PtcLexA promoter for GAL4 expression when exposed to Stit in Drosophila. Wing disk where PtcLexA>GFP is shown as fragments within the pouch and notch. Stit-transformation of cells (Stit>Ptc>GFP) leads to more diluted fragments of GFP-positive cells. The areas of GFP expression are consistent with that of the control.
Stit>Ap>GFP, Stit-transformed cells, showed a smaller area of GFP-positive cells than that of the control. This result was consistent for both samples of Stit>Ap>GFP (Figure 8). The first sample was disturbed due to the positioning on the objective glass, however, there were still GFP-positive cells present within the wing pouch, which were more elongated than the control. The second sample showed a strongly reduced expression region in the pouch, with two separate fragments instead of one larger fragment.
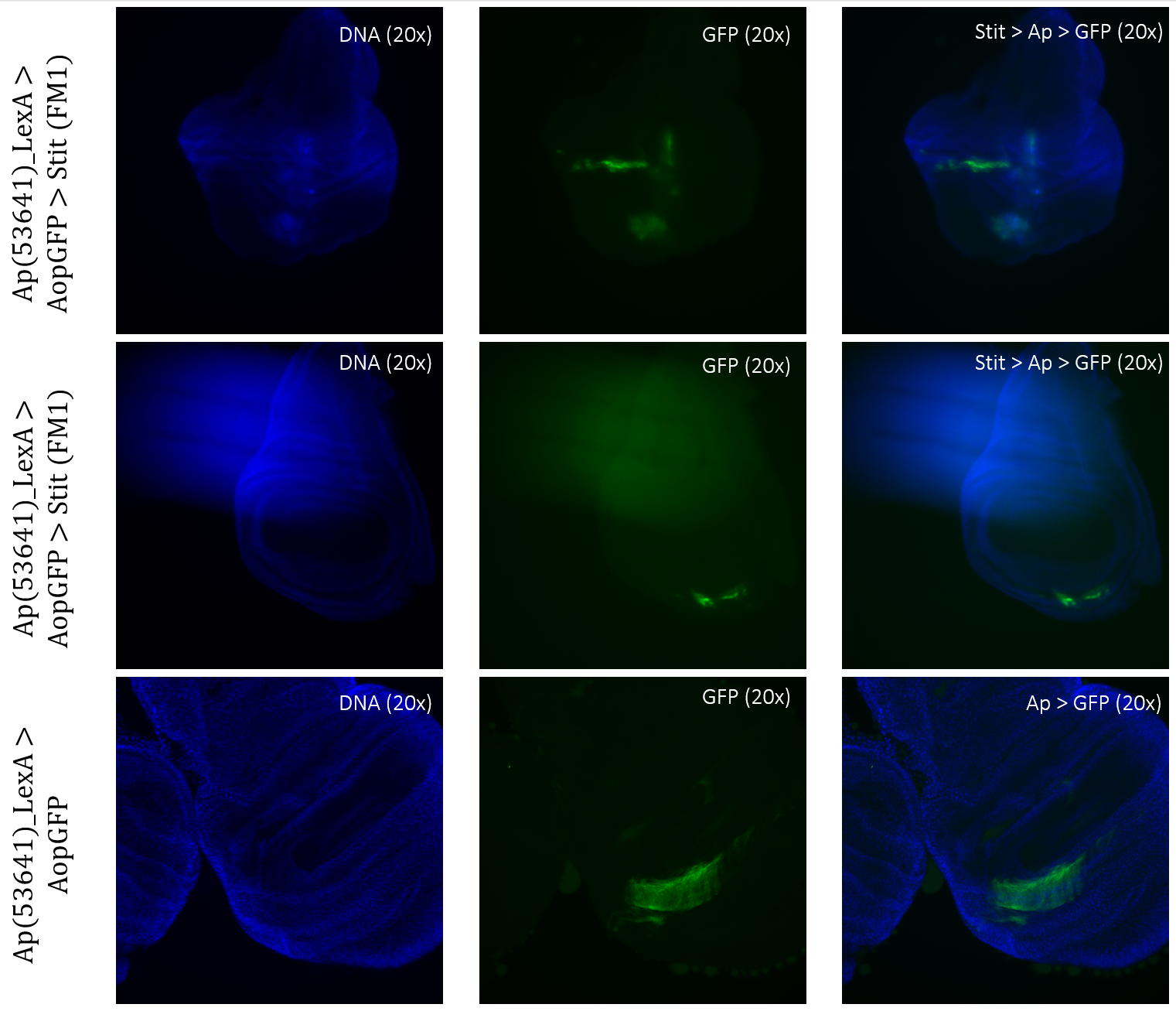
Figure 3.6: Tissue samples from ApLexA promoter for GAL4 expression when exposed to Stit in Drosophila. Wing disk where ApLexA>GFP is shown as fragments within the pouch. Stit-transformation of cells (Stit>Ap>GFP) leads to more diluted fragments of GFP-positive cells within the pouch of the imaginal wing disk. The areas of GFP expression are somewhat consistent with that of the control.
About the author

Photo credits: Aurora Farkas
I am incredibly grateful to my supervisor, Fergal O'Farrell, who has provided me with this opportunity. In terms of both my academic and professional goals, this experience has been a turning point, and I am eager to see where it takes me next.”



1 Response
[…] N. (2023). Characterizing Expression Patterns of LexA and LexAop Lines for the Development of a Drosophila Canc…. Bikuben […]