The Hidden Pollinators of Inner Hardanger: a study in Hoverfly (Syrphidae) Diversity, Abundance, and Phenology

Material & methods
Choice of data analysis
The data analyses for this study included: i) Shannon-Weaver diversity index and ii) species accumulation plots.
The Shannon-Weaver diversity index was used as Alpha Diversity estimate used for the evaluation of species diversity for the three sites. Alpha diversity is an evaluation of how diverse a sample is (Willis, 2019). The Shannon-Weaver diversity index gives a numeric estimate of species diversity and richness for a location (Bobbitt, 2021; Ortiz-Burgos, 2016). The formula for calculating Shannon-Weaver diversity index is:
![]()
where:
pi = proportion of each species in sample
log2pi = natural logarithm of pi proportion
Shannon-Weaver diversity index gives a value between 0-4.5, where the usual values range between 1.5-3.5 (Bobbitt, 2021; Ortiz-Burgos, 2016).
Species accumulation curves were used to estimate species diversity and richness and is a representation of sampling effectiveness for a site (Deng et al., 2015). If the curve of a species accumulation plot is flattened, the samples analysed are representative of the whole species diversity for a site (Deng et al., 2015). If a species accumulation plot is still rising, the data analysed is an underrepresentation of site-specific species diversity, and further data is required to accurately represent all species that are discoverable (Deng et al., 2015). The shaded area of a species accumulation curve shows a 95% confidence interval (Deng et al., 2015).
Sampling design
Triplets of pan traps (yellow, blue, and white) and blue vane traps were placed in the locations Djønno, Urheim and Opedal in Hardanger, Western Norway between late April – mid-June 2022. Djønno and Urheim are located in relatively rural areas, whereas Opedal is located in a more urbanized area. Six triplets of pan traps and three blue vane traps were placed in each of the three locations (Høydal, 2022). Both types of traps contained a soap water mixture to break the surface tension of the water to ensure insects drowned when caught (Høydal, 2022). The insects were sampled through four sampling periods separated by 14 days to prevent over trapping too early in the season (Høydal, 2022). In every sampling period the traps were placed in the field for four days and only emptied every two to three days (due to limitations in available workforce) (Høydal, 2022). Trap contents were bagged and labelled and stored in a freezer prior to identification.
In the laboratory, insects were defrosted and all insects with a false wing vein Vena spuria were selected. After hoverflies were identified to species, they were pinned through thorax with a size 000 pin needle. Individuals identified were given a species_ID. Every individuals’ identification was double checked using materials provided by The University Museum of Bergen, of which included Swedish identification literature and pinned museum examples of the most abundant species identified (Bartsch, 2009b; Bartsch, 2009a; Gammelmo, 2017).
Data Analysis
Data summarised in the excel sheet was exported and loaded into an R Studio workspace where the software R version 4.0.3 was used to process the data (The R Foundation, 2020). To visualise species diversity and abundance, phenological activity and trap colour and trap preference we used the plot() and barplot() functions. The values of Shannon-Weaver diversity index for each of the three sites were calculated and were used as a numerical estimation of diversity. Shannon-Weaver diversity index was calculated using the diversity function, diversity() from the vegan package version 2.5-7 (Oksanen et al., 2022). To check if the sampling portion was an adequate representation of species in the areas, we produced a species accumulation curve. The species accumulation curve was produced by firstly collecting data for species and all traps as a value using the table() function. Secondly, using the default extract method for the specaccum() function on the values sorted into a table, a polygon plot was made by using the plot() function with the specification of ci.type being “polygon”.
Results
Phenological activity
Hoverflies were found to be mostly active during the summer months of June (Figure 1). The highest abundance of hoverflies was found to be mid-June in post-flowering period of apple trees (Figure 1).
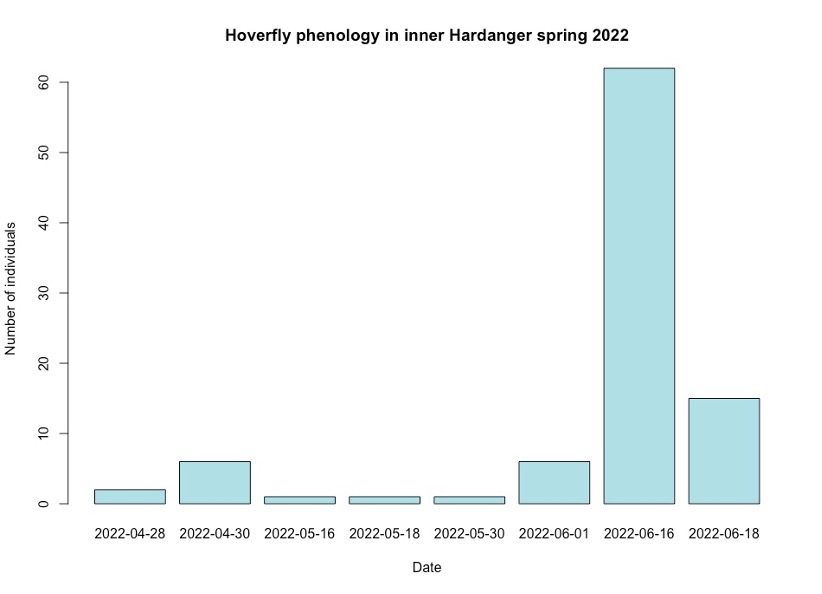
Figure 1: Bar graph showing hoverflies caught on dates throughout the field season in total for all locations.
Shannon-Weaver diversity index
The Shannon-Weaver diversity index for Urheim was approximately 2.66 and was the most diverse site. Close to Urheim on the diversity index was Djønno with an approximate value of 2.52. The least diverse site with an approximate value of 1.71 was Opedal.
Species accumulation curve
The species accumulation curve did not flatten (Figure 2). Due to the rising graph, individuals of the hoverfly family Syrphidae collected was likely an underrepresentation of the available hoverflies in Djønno, Urheim and Opedal (Figure 2). The species accumulation curve did not represent the total species richness for inner Hardanger (Figure 2).
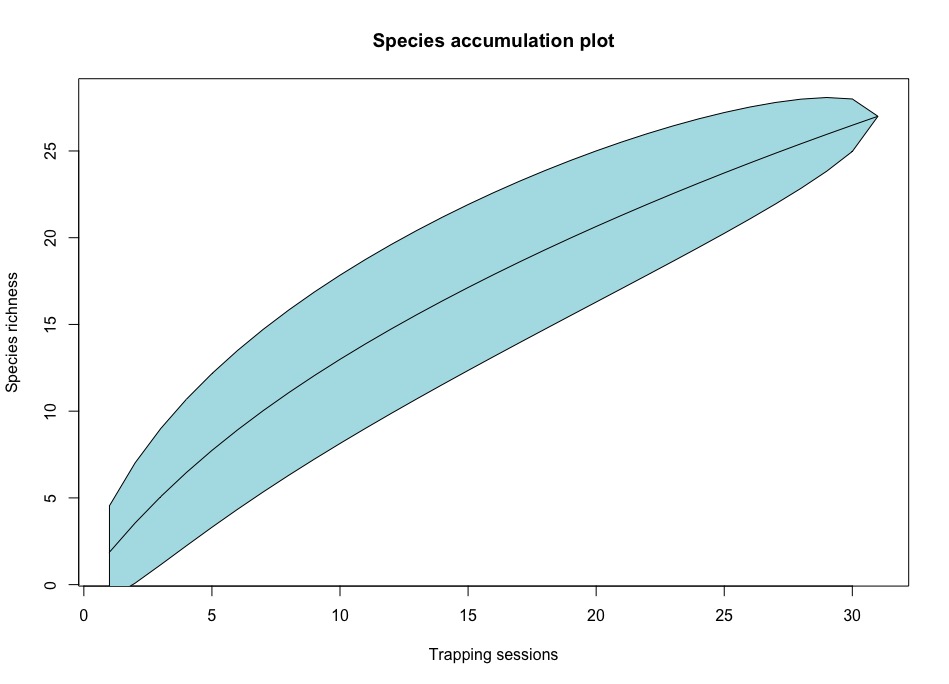
Figure 2: Species accumulation curve for hoverflies caught in all trap types throughout the entire field season.
Species abundance
The most abundant subfamily was found to be Eristalinae, representing 65.96% of all the individuals caught (Figure 3). Syrphinae represents the remaining 34.04% of the individuals caught (Figure 3). There were no individuals caught in the Microdontinae subfamily, and it is therefore not represented in this sampling.
Cheilosia spp. were found to be the most abundant hoverflies, representing 26.6% of all individuals collected. Collectively, Xylota spp. was nearly as abundant as Cheilosia spp., with a representation of 22.34% of individuals.
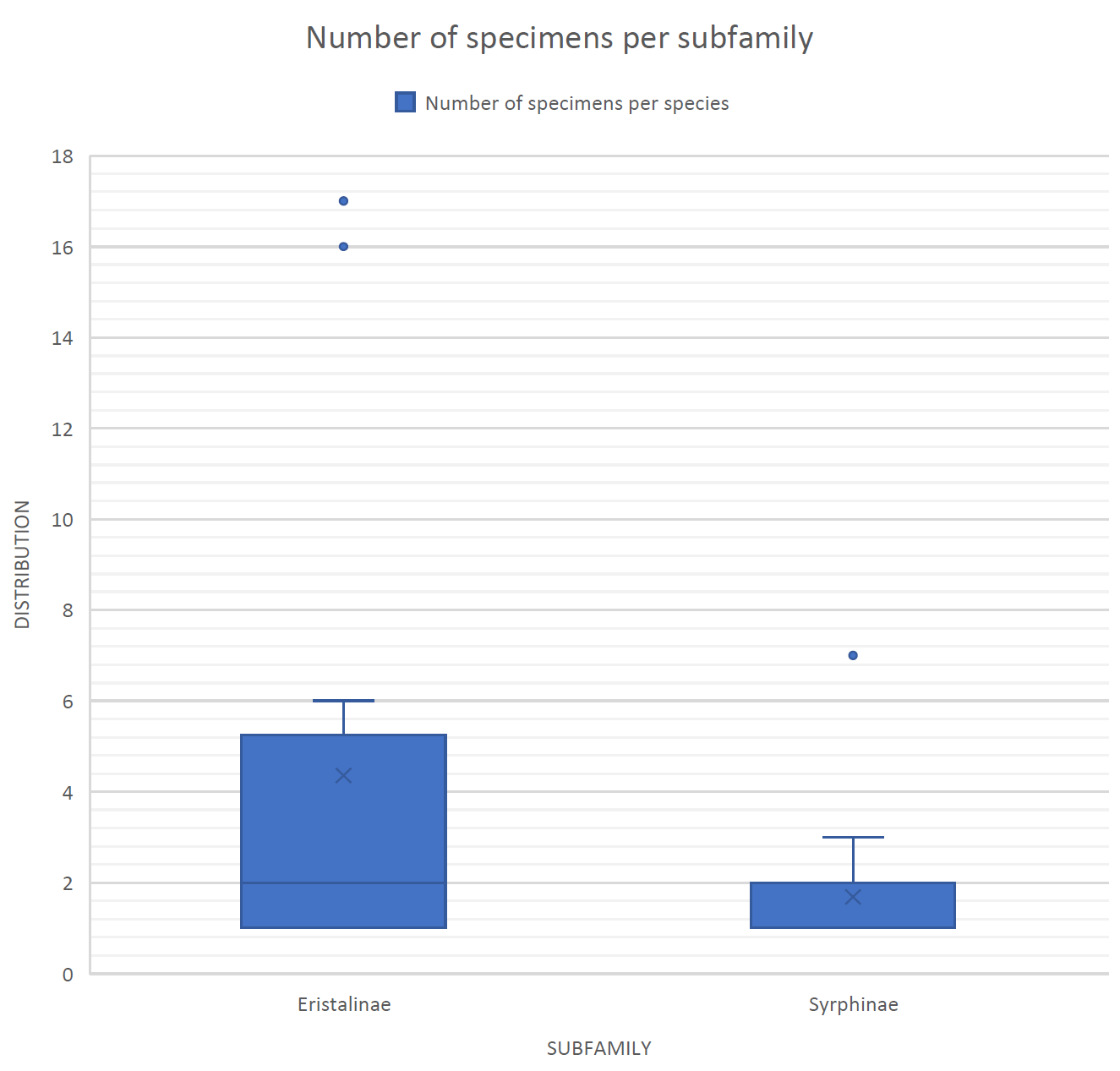
Figure 3: Boxplot displaying the distribution of specimen found for subfamilies Eristalinae and Syrphinae. No data for subfamily Microdontinae.
Site-specific Species abundance
The highest abundances were found in Djønno, with 63 individuals (25 females, 38 males) (Figure 4). Urheim and Opedal were both similar in terms of hoverfly abundance (Figure 4). Identified from Urheim were 15 individuals (6 females, 9 males) (Figure 4). From Opedal, 16 individuals (11 females and 5 males) were identified (Figure 4). A disclaimer is placed here to inform that there were dates with missing trap materials for both Djønno and Opedal.
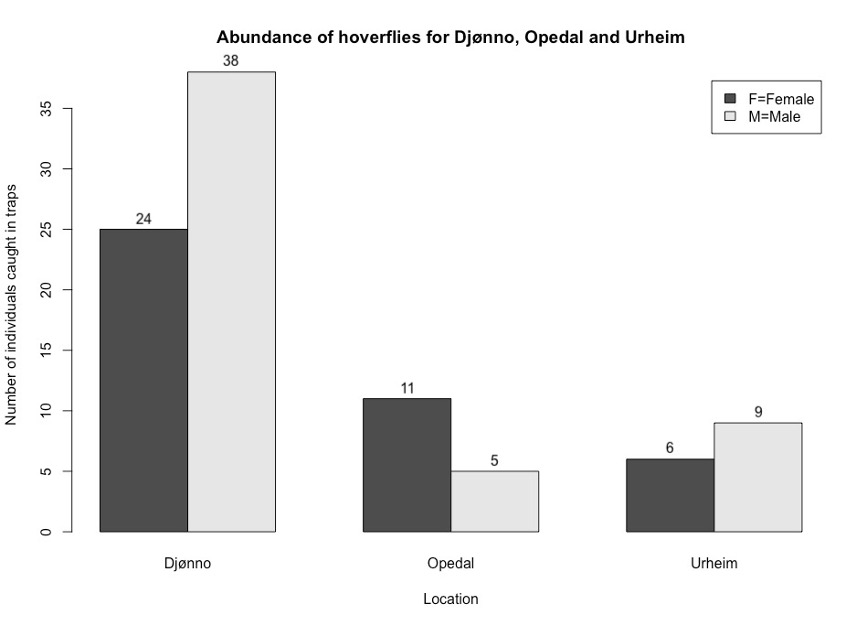
Figure 4: Species abundance for Djønno, Opedal, and Urheim with gender distribution.
Methodological effectiveness
Insects collected using pan traps and vane traps under-sampled the available species for all sites (Figure 5). Pan traps were more effective than vane traps (Figure 5). Yellow pan traps were more effective than white and blue pan traps (Figure 6). 94 individuals of the hoverfly family were identified from this field season.
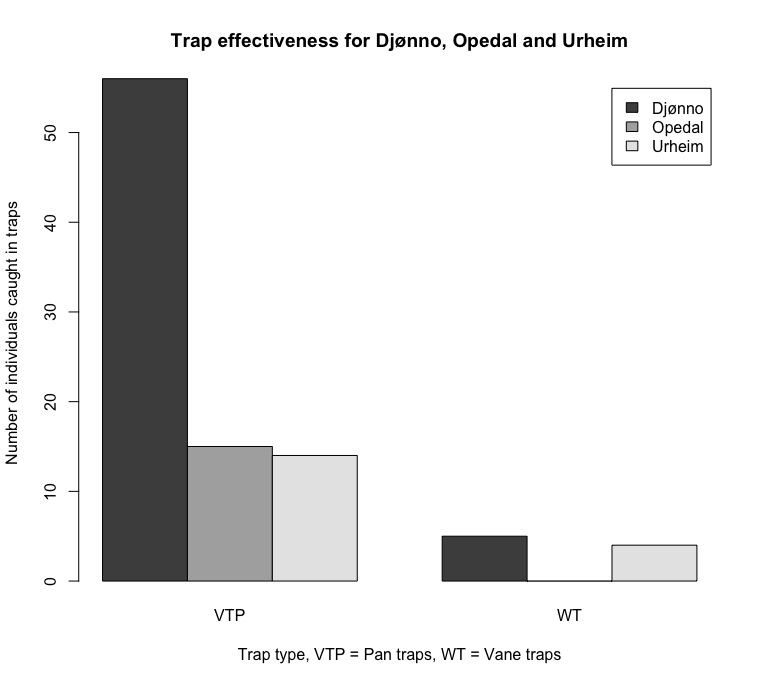
Figure 5: Distribution of individuals captures in 1) pan traps (VTP) and 2) vane traps (WT).
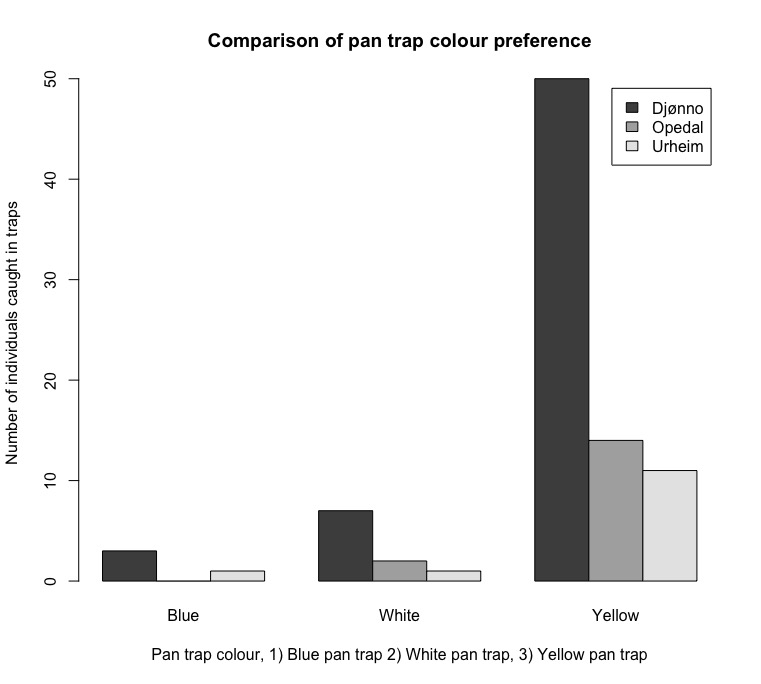
Figure 6: Distribution of individuals per trap colour for all three locations.
About the author

Sara Rodrigues de Miranda
If you had told me before I started my bachelor that I would be the crazy bug person, I would have laughed in your face. Hindsight is 20/20 and entomology has nevertheless become a subject near and dear in my heart. The work that goes into the taxonomy, the puzzle of finding out what species we´re dealing and constantly learning about new aspects of a family of insects is a fundamental part of why I enjoy what I enjoy. I chose to take BIO299 because I had a prior taste for species identification and wanted to further explore practical lab experience with a larger assignment. During my work in BIO299, I unexpectedly fell head-over-heels in love with the hoverfly family, and it has, and will likely remain my passion for quite some time.



{kind=link}
1 Response
[…] de Miranda, S. (2023). The Hidden Pollinators of Inner Hardanger: a study in Hoverfly (Syrphidae) Diversity, Abundance, and…. Bikuben […]