Investigating the number and colour of photophores of Maurolicus muelleri in two Norwegian fjords

Materials & methods
Research Area
The study was carried out between 24–30 September 2022 in two fjords on the west coast of Norway: Masfjorden and Fensfjorden. Masfjorden is a 494m deep, 24km long arm of Fensfjorden, which at its longest point is 50km and 583m deep. These fjords are separated from each other by a 75m deep sill. Data was collected from both fjords to provide the possibility to compare the influence of environmental conditions on the photophores and M. muelleri itself. The vessel used for transportation and data collecting was G. O. Sars, a research vessel shared by the Institute of Marine Research and the University of Bergen.
Field Methods
Individuals of M. muelleri were collected using a pelagic trawl fitted with a multisampler, with three codends at fixed depths (sub-layers) of 300m-200m, 200m-100m, and 100m-0m with an oblique haul through each layer. We trawled for approximately 10 minutes per codend, with an average speed of 2.5-3 knots. The multisampler had a mesh opening of stretch 22mm. Two trawl hauls were done in each fjord and each trawl was done between 8pm and 4am. We took a semi-random subsample of maximum 34 individuals of M. muelleri from each codend. If individuals were damaged to the point where it would have been impossible to count the photophores, they were discarded, and new individuals were selected, based on whether counting the number of photophores appeared possible or not. When the total number of individuals was 34 or below, all individuals were taken. In addition, we collected a random sample of M. muelleri from each codend, to compare with our handpicked individuals showing whether they are representative of the total catch. The subsampled individuals were put on a laminated sheet and scanned with Canon CanoScan LiDE 400. The scans were measured in ImageJ, using standard length. We registered whether the fish had green or pink photophores, and individuals with at least 40% pink photophores were assigned ‘pink’. As individuals had several shades of pink, from pale pink to purple, we simplified their colour to only pink, reducing errors due to human subjectivity.
Lab Methods
In the lab, we defrosted and weighed each fish individually (wet weight) in its plastic bag and subtracted the average weight of the bags. We then registered the number of photophores for each section of the fish, following the protocol described by Sutton et al. 2020 (p. 68) for the first 25 individuals of each codend. In the case of individuals being very damaged, we discarded that fish and used one of the remaining 9 individuals. After counting photophores, we extracted sagittal otoliths to determine age. We determined the age by counting increments on the otoliths, starting at age 0, with no, or a very small increment and assuming formation of the rings begins at hatching and continues until the adult stage. Some fish over 1 year were described as 1+ (written Age 1.5), due to the outer winter ring being the same size as the outer summer ring. Age 2 had two clear winter rings. The data was processed in RStudio, R version 4.2.1 (2022-06-23). We used data collected from the same field course, conducted in previous years and in addition to the data collected this year on size distribution for M. muelleri, we checked whether our semi-random sample was a good representation of the population.
We made two models in R, one linear model describing the total number of photophores, with total number of ventral photophores as the response variable, and the fjord, depth, body condition and age as the predictor variables. The second model was a binomial model which used photophore colour as the response variable with the same predictor variables as the first model. Both models were made by finding the best fit through removing parameters in order of least significance.
An ANOVA was performed to check for significant relationships between the different parameters. This was repeated for other comparative analyses. Post hoc pairwise comparisons were also made to further investigate significance found in the ANOVA tests performed.
Body condition was calculated using Fulton’s body condition factor K (Fulton, 1904), 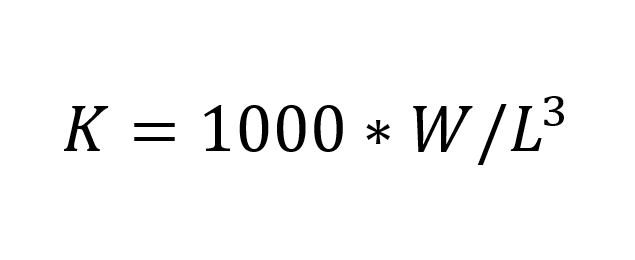 (I)
(I)
where W is weight (g), and L is length (mm). 1000 is a scaling factor. This condition factor was used to compare against other measurable parameters, rather than estimating the body condition itself. In total, we weighed, registered colour, and counted photophores of 258 individuals, 138 from Fensfjorden and 120 from Masfjorden. 75 fish were not counted, only weighed and colour registered. During our analysis, we found that the number of photophores on the head was constant on all individuals. The only notable variation in photophore number came from the ventral section on the fish (PV, VAV, AC, as given by Sutton et al. 2020, p. 68). By only analysing these photophores, we can include a larger number of fish due to ignoring the instances where it was not possible to record the photophore number. In total, we have 192 individuals with counted ventral photophores, 120 from Fensfjorden and 72 from Masfjorden.
Results
The comparison between the length distribution of our data and the total length distribution of M. muelleri caught during night-time on the cruise showed our samples represented the total range of lengths.
Photophore Number
Only 124 of the counted individuals were in a condition where we could count all photophores. The mean number of total photophores for these individuals were 138 ± 2.61. Of all counted individuals, we were able to fully count the ventral photophores of 225 (Table 1). Of these, 92 where from 0-100m, 93 from 100-200m, and 40 from 200-300m depths. The mean number of ventral photophores for these fish were 87.9 ± 2.62.

Table 1: Depth distribution of individuals of Maurolicus muelleri used in the analysis of numbers of photophores. Numbers from both fjords and total count are included.
Most individuals sampled in the study were of age class 1 (n = 202), with much fewer samples from age class 0 (n = 2), age class 1.5 (n = 15), and age class 2 (n = 6).
There was no significant variation in photophore number related to body condition (ANOVA; F(1, 221) = [0.83], p = 0.36), between the two fjords (ANOVA; F(1, 220) = [0.39], p = 0.53), or with age (ANOVA; F(2, 217) = [0.35], p = 0.412). During model selection, we found the only explanatory variable to be depth (p = 0.02), so further downstream analyses were performed using this best fitted model. We found a significant difference in number of photophores in relation to depth (ANOVA; F(2, 222) = [3.78], p = 0.02), with the mean number of ventral photophores of individuals at each depth being: 0-100m = 88.4 ± 2.67, 100-200m = 87.7 ± 2.50, 200-300m = 87.1 ± 2.59.
A post hoc pairwise test between total number of ventral photophores and individual codends showed no significant results between the depth layers 0-100m and 100-200m (Post hoc; t ratio = -1.85, estimate = -0.70, df = 222, p = 0.16), or 100-200m and 200-300m (Post hoc; t ratio = -1.16, estimate = -0.57, df = 222, p = 0.48). There was, however, a statistically significant result between the shallowest 0-100m and deepest 200-300m layer (Post hoc; t ratio = -2.59, estimate = -1.27, df = 222, p = 0.03).
In total, there was a trend of higher number of total ventral photophores in the shallowest layer, but no trends or relationships between age, body condition or fjord (Figure 1).
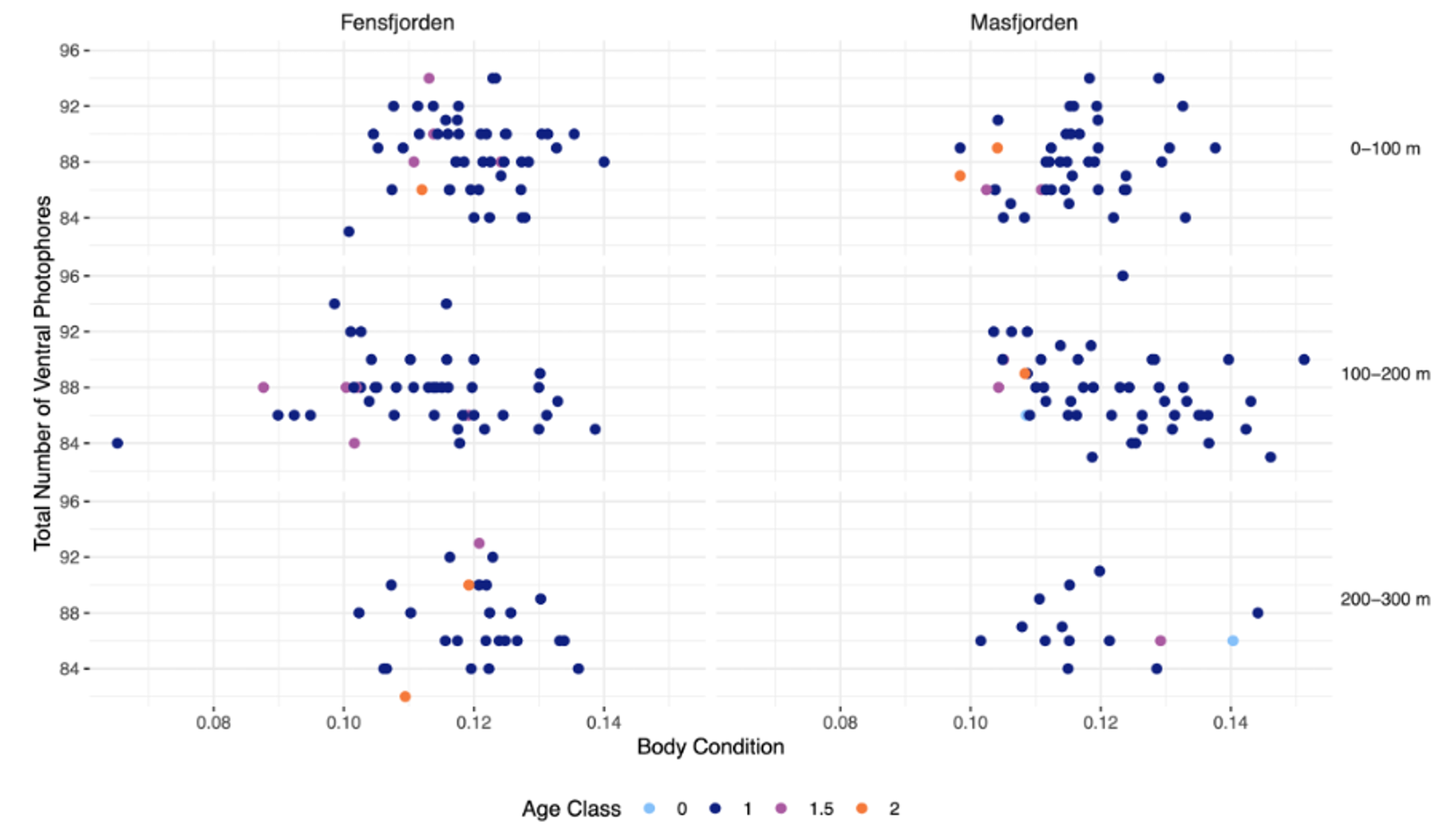
Figure 1: Number of ventral photophores in relation to body condition, depth, and fjord. The relationship between body condition and number of ventral photophores of Maurolicus muelleri in the two fjords Fensfjorden and Masfjorden. The depth at which the M. muelleri where caught are also included. The age of individuals is shown by the distinct colours of the points. Age was decided by counting otolith increments.
Photophore Colour
In total, we registered the colour of 289 individuals from all depth layers (Table 2). 217 of the individuals were green (75%) and 72 were pink (25%).
182 of the counted individuals came from Fensfjorden; 143 were green (78%), and 39 were pink (22%). The remaining 107 were from Masfjorden, of which 74 were green (69%) and 33 (31%) pink.

Table 2: Depth distribution of individuals of Maurolicus muelleri used in the colour analyses. Numbers from both fjords and total count are included.
Green coloured photophores occurred in higher numbers in both fjords and depths, except for the deepest depth where the proportion of pink is greater (Figure 2).
Using the best fitted model including only depth and fjord, a chi-squared test showed significant relationships between the colour distribution and fjord (χ2 (1, N = 289) = 16.67, p = <0.001), depth layers (χ2 (2, N = 289) = 44.50, p = <0.001). Age (χ2 (1, N = 289) = 2.11, p = <0.15) and body condition (χ2 (1, N = 289) =1.91, p = <0.17) showed no significant effect on the photophore colour.
A post hoc pairwise test between the observed colour of ventral photophores, and the depths sampled showed a significant difference in colour between the deepest (200m-300m) and middle layer (100m-200m) (Post hoc; p = <0.001 z ratio = 4.71, estimate = 1.92), and between the shallowest (0m-100m) and the deepest (Post hoc; p = <0.001, z ratio = 6.11, estimate = 2.73). There was, however, no significant difference between the shallowest and the middle layers (Post hoc; p = 0.06, z ratio = 2.24, estimate = 0.81).
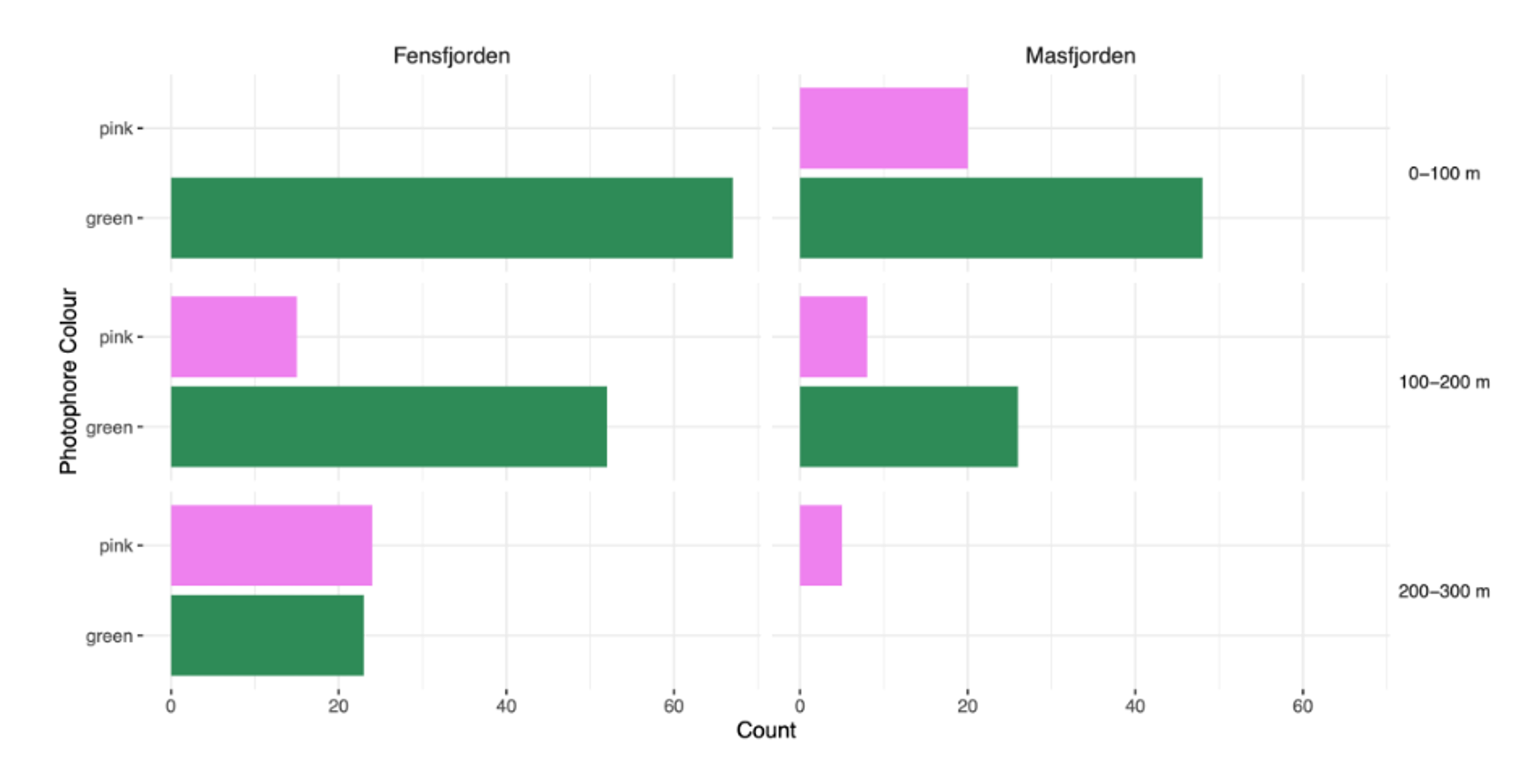
Figure 2: Count of photophore colour in relation to depth and fjord. The number of Maurolicus muelleri with green and pink photophores in the two fjords Fensfjorden and Masfjorden. The depth at which M. muelleri were caught is also included.
About the authors

The “Fjord Bros”
Hi! We are the “Fjord Bros”. A group of MSc marine biology students, but more importantly - a group of friends. The Fjord Bros consists of Anette Aune (ecology and ethology girl), Joe Lawrence (genetics guy), Kristian Fjeld (mesopelagic fan), and Adam Mortensen (fisheries man).
Our group was formed by random assignment during the BIO325 “Ocean Science” course in 2022, and as fate would have it, through our shared passion and curiosity for the oceans - especially the mesopelagic fjord ecosystems, we developed a strong bond and quickly chose to name ourselves the Fjord Bros.
Our paper all started with a couple of very simple questions. How does the number of light organs (photophores) change as the mesopelagic fish “pearlside” (Maurolicus muelleri) grows? And why do some individuals have pink light organs, as opposed to green? To answer this question, we spent a week aboard G.O. Sars in Masfjorden and Fensfjorden, working long shifts at the wet lab sampling hundreds of pearlsides caught by trawling.
Back on land, countless hours and late evenings were spent counting each individual fish in a microscope. Through thick and thin, we stood together, determined to find the answers. As a result, through all this hard work, you are now able to read our paper which we are very proud to present. We hope you enjoy it.
During our writing process, we had regular meetups at K1/K2, to feast on the waffles served by biORAKEL. We would like to extend our gratitude to the people working there, ensuring that we had a great environment to meet and work in.


