The proximate mechanisms behind fear: an IBM approach to understanding behavioral changes in songbirds and the subsequent evolutionary trajectory

Materials & methods
The architecture of our model is depicted through Figure 1. For each iteration, aka generation, the digital birds are sent into an array consisting of 100 cells along the x-axis. Each cell represents a habitat consisting of two values: food availability and risk of predation. These values are randomly generated so that each cell/habitat is unique. The birds “fly” at random to a cell. The actions of the bird upon landing at a specific cell are dictated by both their gene, and how their emotional state is at that moment. If they are sufficiently afraid, they will not eat. If they are sufficiently hungry, they will not be afraid. This can either aid them in gaining mass or harm them upon meeting a predator. If they are too hungry to mind the risk of themselves being killed by the predator, they are at risk of being killed. The phenotypic range of their actions is constricted by their genetic value, which is randomly sorted throughout the population at the beginning of each simulation. Therefore, there will be a genetic diversity in place and selection of an advantageous gene will be possible.
In our model, we used elements from the theory of the global organismic state (GOS) (LeDoux, 2012) in the following ways: If our bird is beyond an emotional threshold where it becomes fully fearful, the bird will stay put and not fly from cell to cell in search for food. After a while, it will grow hungrier, and fear will reduce to a point below the threshold. Mathematically, this can be expressed in the following way:
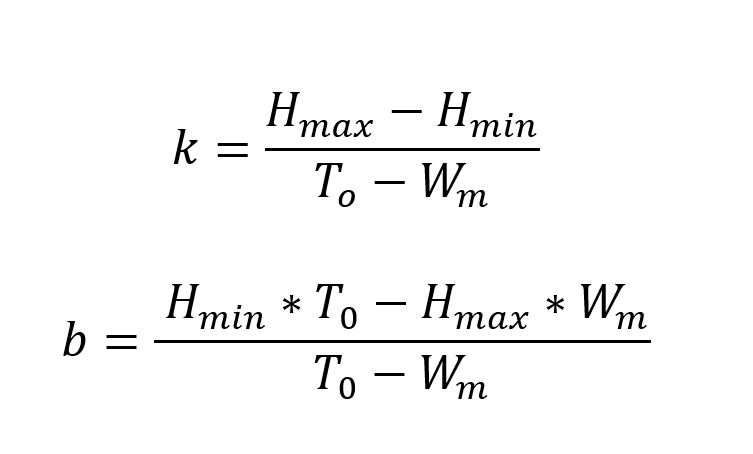
Here, k represents the slope defining the temporal aspect of the transition between fear and hunger, and b is the intersection point of which the birds’ emotional state is at a certain point in time. Hmax and Hmin refers to “maximum hunger” and “minimum hunger” that the bird experiences. T0 is the weight-threshold of the birds and is set to -25% of the initial weight of the digital songbird. If a bird’s mass drops below this threshold, it is considered dead from starvation. Wm on the other hand, gives the value for the birds’ maximum mass, which in our model is 25% larger than its initial starting weight. We have not considered the weight of chicks since we, in our model, assume all birds to be adults.
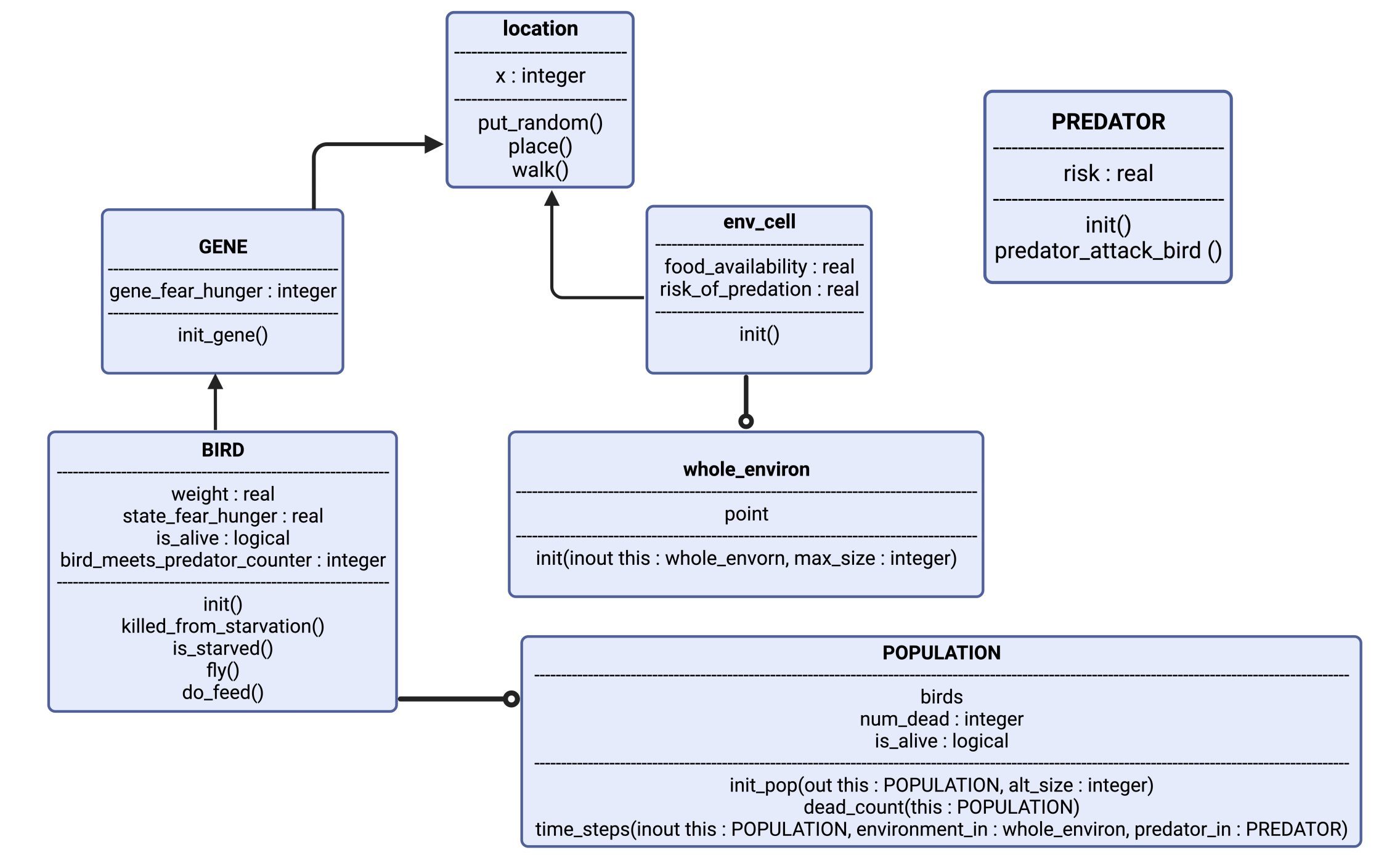
Figure 1: Visualizing the model: The object “BIRD” has attributes weight, state_fear_hunger, is_alive and a counter for when it encounters a predator. It has certain actions as well, such as: is_starved and fly. “BIRD” is controlled by “GENE”, which gives it the basis of its phenotype. “BIRD” has now a genetically defined personality, and expresses this in “location”, which is the exact point in the “env_cell” (short for environment of the cell), which is a part of “whole_environ” (the entire environment consisting of 100 cells, i.e., habitats.). In each cell there are also a few instances of “PREDATOR”, which can spawn randomly based on parameter-set probability. Each simulation spawns several thousand “BIRD”, which gives us the “POPULATION”.
Object-oriented modeling
The model consists of “objects” (figure 1). Each of the objects are representations of agents in our model, or the environment the agents act within. The agents are the songbirds (BIRD) and the predators (PREDATOR). Each of the objects have their own characteristics that define what qualities the objects possess; where the objects are (i.e., which habitat is the bird in), and how the objects’ interactions affect their state. The qualities mentioned are assigned values set by parameters in the code. An example is the object “BIRD”, which has the real value of “weight”. In the code, we set the initial weight to “20.0” (grams), which will be the initial value of the birds every time we initialize the code. By using an object-oriented model, we can better mimic the encounters a bird might have with food and risk in nature.
The Genetic Algorithm
In our model, we also created a genetic algorithm (GA) as stated by Holland (1992). When each of our objects: "Environment", "Predator" and "Parent population" (Figure 2), are initialized and iterations for each generation are done, the GA sorts the songbirds and calculates their fitness". It does so in the way illustrated by the smaller circle in Figure 2: each parent population undergoes a series of timesteps, which gives the order of their actions. After the songbirds’ actions are completed, the fitness of the remaining songbirds is calculated based on their gained mass. Of all the songbirds, 25% of the fittest are automatically chosen for reproduction by the code. To further genetic variation in the offspring generation, we have also included an additional 25% chosen at random from the remaining population. Therefore, the next generation in our model will have genes from these 50%. This process is shown in Figure 2 as “select_reproducing”. After this process of selection based on fitness, we submit our new genes to mutation. According to the literature, we chose the parameter for the mutation probability rate to be 4.6 × 10−9 (Smeds et al., 2016). The selection with added mutation results in the parent populations´ offspring (“offspring_population” in Figure 2). The offspring will become the main population for the next generation, completing our single iteration of the GA.
In our model, we chose 100 iterations of the GA, meaning we ran the simulation for 100 generations of songbirds. Since the evolution of a single gene was shown, we believe this number of generations should be sufficient. We fully acknowledge and stress the fact that this is not reality. It is however sufficient to prove that our model works and that evolution happens.
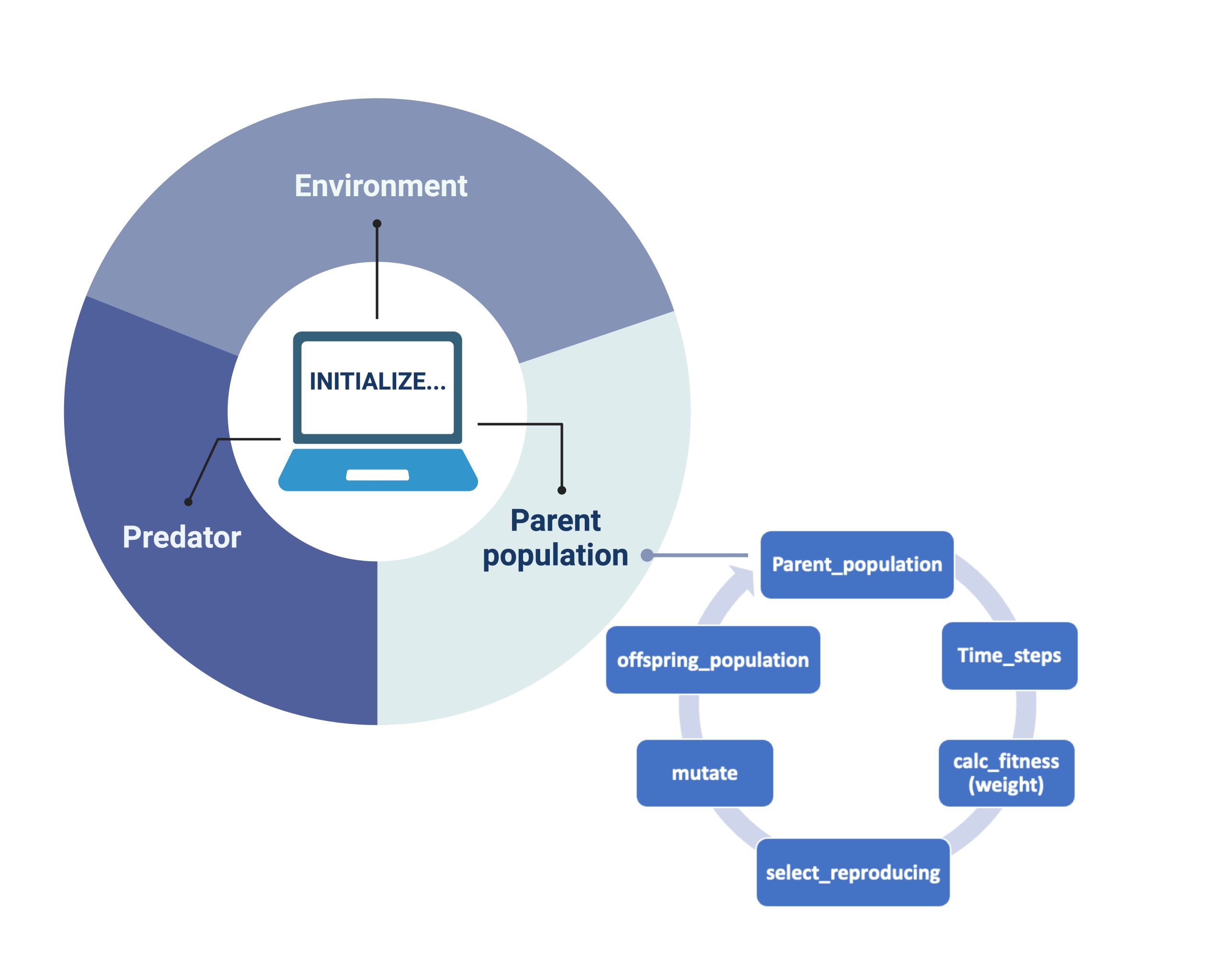
Figure 2: How the model works. For each Initialized simulation our objects (the parent population of birds, the predator and the environment interact). For the birds that show the greatest fitness (measured only by “best mass”), a selection undergoes mating and creates the offspring generation. To incorporate genetic diversity, we also select a certain number of birds that are less fit than optimal and introduce mutation of genome in each generation.
Modeling tools
Fortran (Formula Translation)
Fortran is a widely used programming language in the natural sciences, including biology. Fortran's proficiency in handling numerical calculations and array operations makes it suitable for modeling natural events and data analysis. The language provides a broad spectrum of mathematical functions and supports advanced operations, enabling biologists to develop complex models that can capture the dynamics of biological systems.
SVN (Subversion)
SVN is a version control system used in software development. With SVN, developers can work simultaneously on the same code without conflicts, as the system tracks and manages the merging of changes. It allows users to check out a working copy of the project, make modifications, and then commit those changes back to the repository. SVN maintains a comprehensive record of all changes, making it easy to roll back to previous versions if needed.
Visual Studio and Visual Studio Code
Visual Studio is a widely used integrated development environment (IDE) created by Microsoft. It provides a comprehensive set of tools and features for building a variety of software applications, including desktop, web, mobile, and cloud-based applications. Visual Studio Code is a versatile and lightweight source code editor that offers a wide range of features and customization options. Its ease of use, extensive extension ecosystem, built-in Git integration, debugging support, and productivity-enhancing tools make it a popular choice among developers for various programming languages and platforms.
The Code
The code in its entirety can be found and examined at GitHub: https://github.com/Kaw-Han/songbird-evo-model-FORTRAN. The model code can be examined, tested, run, and furthered under the restrictions of the assigned trademark.
Results & Discussion
Our findings indicate that evolution occurs rapidly in our model (Figure 3). The phenotypic expression associated with the gene in our digital songbirds is characterized by moderately cautious behavior during their search for food. This suggests that a moderate level of fear serves as a beneficial trait for increasing fitness, measured by the songbirds’ gain in mass. Figure 4a further supports this result. The population of songbirds experiences a significant decline in the first five generations, followed by a subsequent rise and stabilization. This pattern indicates the presence of selection, with the gene favored by the environment quickly dominating the population. Additionally, the graph in Figure 4c depicting the average mass of our birds contributes to our findings. It shows that the rapid growth observed in the first five generations is not sustainable; suggesting that the less fearful songbirds, which gained mass due to bold behavior, were not favored by the environment.
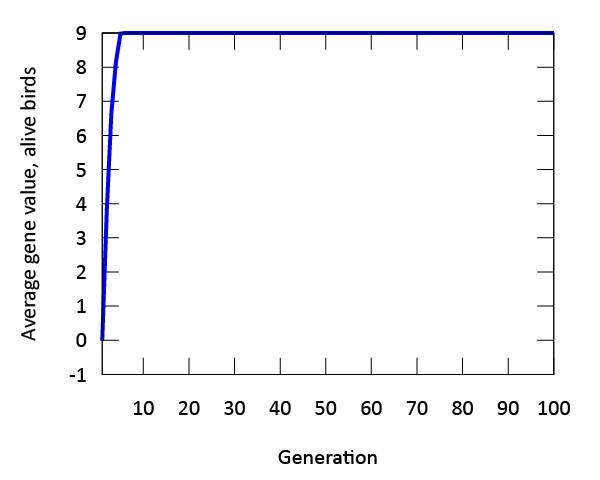
Figure 3: The genetic value "9" is evolutionary stable after approximately five generations. The phenotype expressed by this gene value is associated with moderate amounts of fear.
This point is further supported by comparing this graph to the graph in Figure 4d, which depicts the mass gain of the fittest birds. Unlike the initial increase seen in the first five generations, the graph shows a steady, linear growth rate. This indicates that the most fit songbirds are those expressing the phenotype of moderate fear (Figure 3) from the beginning of our simulation. Furthermore, the dominance of the gene is demonstrated by the standard deviation of mass in Figure 4b. As the songbirds adapt their foraging strategies through selection, the gain in mass becomes almost uniform across the population after five generations.
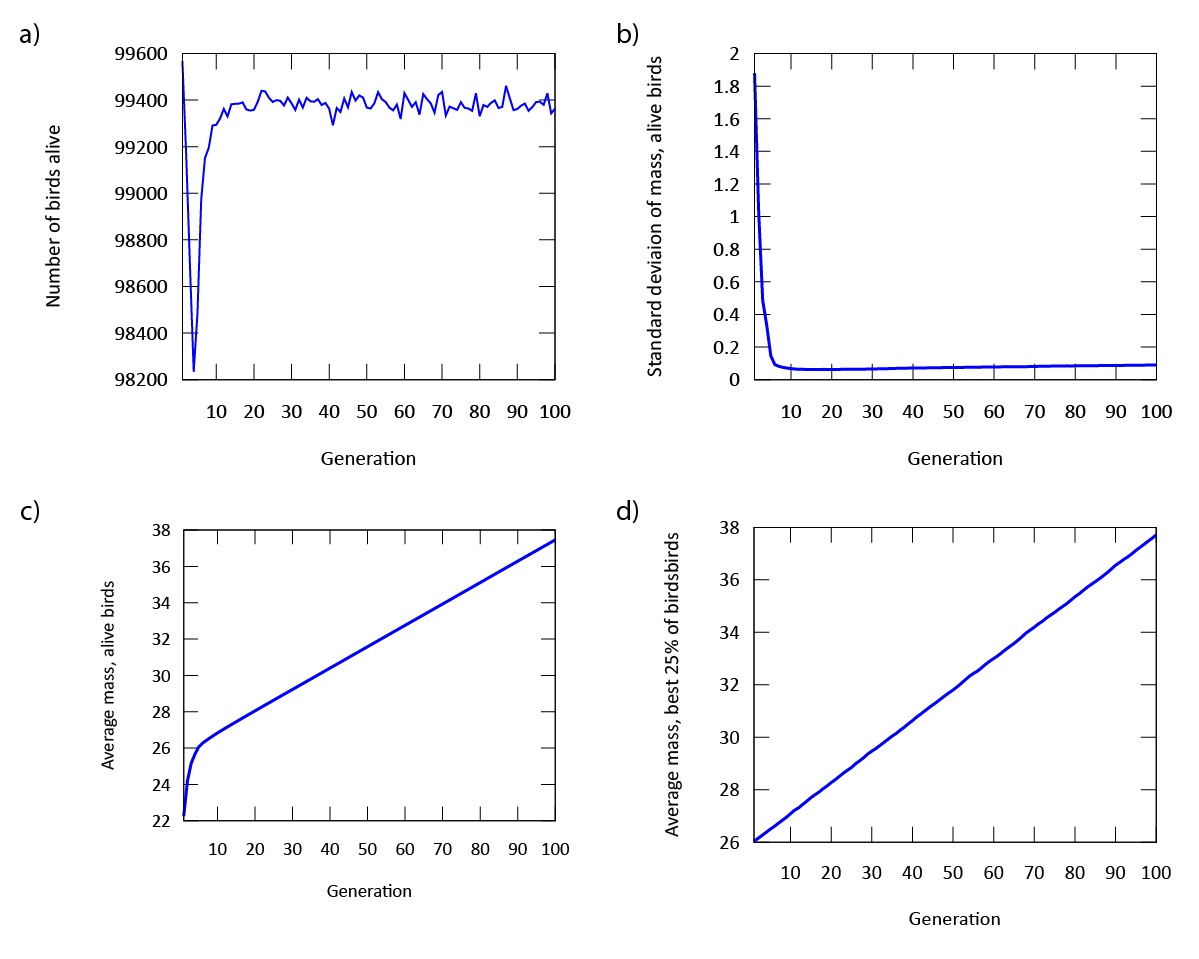
Figure 4: a) The population declines, but quickly regains its numbers after approximately five generations. b) The standard deviation of mass in our population is stabilized after approximately five generations. c) The average mass of all digital songbirds rises significantly throughout the simulation. d) The average mass of the birds with the highest fitness rises linearly and significantly throughout the simulation.
Another important finding in our model is the expression of a single gene through multiple phenotypes. When designing the digital birds with a single gene, we observed variations in the expression of this gene across the population. This plasticity, though constrained by the gene, was evident in our initial population. As time progressed in our model, evolution occurred, and the phenotypic expression we referred to as "moderate fear" remained the main phenotype. This finding is further supported by both Figures 3 and 4, which demonstrate that the fittest birds were those expressing this phenotype from the outset of our simulations.
Through our findings, we present a demonstration of how heuristics (Hutchinson & Gigerenzer, 2005) come into play. The digital birds in our study exhibited responses to predators or hunger based on the information they sensed from their environment while operating within the constraints imposed by their proximate genetic architecture. Notably, we observed variations in the responses of individuals, suggesting that the rules of thumb guiding their behavior may have limitations but are not entirely genetically predetermined. Expanding on this line of thinking, we can question how the concept of the phenotypic gambit (Grafen, 1984, 1991) explains this phenomenon. Although the gene is expressed initially in different ways, the scope of the diversity in phenotypes where still genetically limited. The phenotypic gambit does not consider that the proximate mechanisms sometimes constrain the expression of adaptive behavior, as pointed out by Fawcett et al. (2013) and is therefore sometimes wrong. The numerical findings we present add to the arguments against the simplicity of the gambit and give us cause to further investigate this phenomenon and continue questioning the benefit of the gambit’s exclusion of genetics.
While our model provides only a partial explanation, it offers valuable insights into the findings of Allen et al. (2022). In their study, the songbirds exhibited not only a significant decline in fitness but also a decrease in population size. It is worth noting that the intervention implemented in their experiment was discontinued after five generations. Interestingly, our model demonstrated a similar timeframe before the beneficial phenotype became dominant. On one hand, the fact that the model mirrored real-life events reinforces the validity and utility of the models. On the other hand, it underscores the growing significance of modeling interventions in nature as initial steps toward comprehending population dynamics and the effects of factors like predation in novel environments.
It is important to acknowledge the immediate issues raised by this thought experiment. Firstly, the ethical implications of prolonging or conducting similar experiments as Allen and co-authors (2022) on free-living populations would not align with the standards of conduct in biological science, as recognized by the ASAB Ethical Committee/ABS Animal Care Committee (2023). Secondly, adhering to animal welfare guidelines prompts further research inquiries, such as investigating the impact of invasive urbanization on wildlife inhabiting forest areas.
This underscores the growing importance of utilizing computational biology to develop models that aid in predicting likely scenarios. While models heavily rely on data obtained from field studies, they offer possibilities to explore nuances, address knowledge gaps, and even challenge established theories without sacrificing lives or causing harm to nature. In our model, we could simply design the neural capacity of the songbirds, their habitat, their physiological needs, and their psychology. By applying what we already know, we can provide information valuable for future studies. The need for such an application of our method, is best expressed by the 3Rs (ASAB Ethical Committee/ABS Animal Care Committee, 2023; Sneddon et al., 2017): Replacement, reduction and refinement. Through models and simulations, we can replace real animals with digital ones. We are then able to reduce the number of animals affected. Finally, we can refine future studies by using predictive modeling to suggest ideal points of research that affects animal life in the future.
Expanding the code
We fully acknowledge the simplicity of the model. The fitness of our birds is measured solely by their gain in mass. Although this represents successful strategies, more modules, subroutines, and functions could be interesting to include. Alternate strategies for survival and reproduction are commonly seen in nature. This would provide an interesting addition to our model. We could introduce more genes, more environmental factors such as seasonal changes or migration into the population.
In the times of environmental change, we also suggest introducing objects that represent installations placed in nature by humans. Windmills are such an object and are by many considered as intrusive to nature. Data collected from the existing sites containing windmills, can be utilized to create a valuable addition to our model, and subsequently to stake-holders on all sides of the matter.
Summary
In this article, we utilized an individual-based model (IBM) to investigate the proximate mechanisms underlying the response patterns exhibited by songbirds, as studied by Allen et al. (2022). While our understanding of the «whole picture» remains incomplete, our model offers valuable insights into how fear-based behaviors contribute to evolution. Firstly, our model demonstrated the occurrence of evolution. Our results revealed that the digital songbirds experienced an increase in body mass, leading to a higher probability of survival and reproduction. We deduced that the fittest individuals possessed phenotypes that provided advantages right from the start of our simulations. The remaining population adopted similar phenotypes within a mere few generations. Specifically, the most advantageous phenotype expressed a moderate level of fear during foraging. Secondly, our model demonstrated that multiple phenotypes could originate from the same genotype. This finding is particularly intriguing when compared to the concept known as "the phenotypic gambit" (Grafen, 1984, 1991). Although our model, like the gambit and previous models, simplifies for the sake of optimization, our findings suggest that proximate mechanisms warrant further attention from behavioral ecologists who favor the gambit's premises in their research. Finally, we propose the need for future development of our model in order to introduce more true-to-nature complexity, and therefore increase the value and precision of our simulations.
About the authors
 Hello and thank you for looking at my paper. My name is Hanif Kawousi and I am currently working on my master’s thesis at the Department of Biological Sciences at the University of Bergen. I believe some of the most interesting questions one could ask are within the field of evolutionary ecology. To answer these questions I am working on developing models that can simplify complex systems in real-life nature into something we can measure, handle and research, without compromising the systems’ integrity itself.
Hello and thank you for looking at my paper. My name is Hanif Kawousi and I am currently working on my master’s thesis at the Department of Biological Sciences at the University of Bergen. I believe some of the most interesting questions one could ask are within the field of evolutionary ecology. To answer these questions I am working on developing models that can simplify complex systems in real-life nature into something we can measure, handle and research, without compromising the systems’ integrity itself.
I would like to take the opportunity to thank my supervisors and co-authors: Sergey Budaev and Jarl Giske, both at Theoretical Ecology Group at UiB. I would also like to thank my wife, Jenny, for all her continuous love and support.


