Let’s talk about sex: Exploring evolutionary explanations for the persistence of sexual reproduction in eukaryotes

2. Discussion
Initially, some quick definitions are in order. Reproduction is the process by which organisms give rise to offspring. Sexual reproduction can be defined as when a new organism is produced by the mixing of the genomes of two organisms of different types (parents) (Reece et al., 2014). Both parents produce gametes through meiosis, during which the chromosomes undergo recombination where genetic information is exchanged between the homologous chromosomes. Through this process every individual gamete produced by an organism are genetically unique to each other. Through outcrossing and the fusion of sex cells, 50 percent of the genetic material from each parent is combined in a zygote. This results in offspring that are genetically different from each other and from their parents. In short, sex is recombination and outcrossing, and the mixture of genes can be considered its primary feature. It does so systematically across the entire genome, breaking down previous combinations and generating new ones. But why?
2.1 The Vicar of Bray and Selective Interference
An early answer to the question of why sex persists dates back to the 19th century and the previously mentioned August Weismann. He suggested that the key benefit of sex is that it generates greater variation for natural selection to act upon, claiming it as "the source of individual variability, furnishing material for the operation of natural selection" (Weismann, 1889). Weismann further argued that sex was just as likely to generate detrimental as beneficial combination of genes, meaning that there is no net advantage to sex for the individuals in any given generation. The population as a whole, however, does benefit, as detrimental combinations are weeded out of the population by means of natural selection, leaving the favorable combinations. Bell (1982) later dubbed Weismann's view on the evolutionary purpose of sex the Vicar of Bray hypothesis. Closely linked to this is the idea that sex reduces selective interference, separating alleles from their genomic backgrounds allowing for more efficient adaptive selection processes (Futuyma and Kirkpatrick, 2017).
It might be important to emphasize that sex does not itself introduce new variation to a population in terms of new genes or alleles. Mutation (or gene flow) is needed for this. Without this, sex only generates new combinations of existing genes, eliminating disadvantageous variants along the way. Without the introduction of new variants, sex actually restricts variation.
After Weismann, a continuation of the Vicar of Bray hypothesis was formulated in terms of population genetics by Fisher (1930) and Muller (1932), among other things incorporating the role of mutations in a clearer way.
Figure 1 illustrates the resulting Fischer-Muller model, which demonstrates an advantage to sexual reproduction through the avoidance of clonal interference. Consider the outcome in an asexually reproducing population if advantageous mutations A and B appear (at different loci) in different individuals. Due to the mutations being beneficial, the clones of these individuals proliferate. Becoming common in the population, the two clonal lines start to compete, potentially driving one of the lineages to extinction (in this case aB). The only way for an AB genotype to get fixed in the population, is for both mutations to independently arise in the same clonal lineage, an unlikely event. In short, clonal interference is a hindrance to adaptive evolution in the population.
For a sexually reproducing population on the other hand, novel advantageous genotypes can be created more rapidly. If allele A and B arise independently in different individuals, the two alleles can rapidly be recombined in the same organism. Sexual reproduction allows the beneficial AB genotype to quickly spread to fixation.
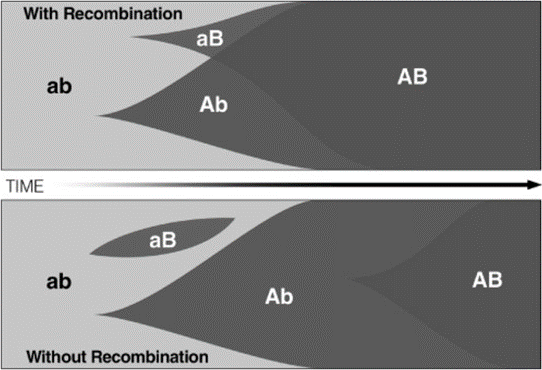
Figure 1: The spread of advantageous mutations in a sexual (top) and an asexual population (bottom). In the sexually reproducing population, the advantageous mutations (A and B) are quickly combined and the AB genotype brought to fixation due to recombination and selection. Without sex however, A can only spread at the expense of B and vice versa, unless both mutations arise in the same clonal lineage.
Crow and Kimura (1965) later provided a modern demonstration of the Fischer-Muller model. Through mathematical formulations they demonstrated the relative rates of incorporation of the new mutations with and without recombination. They found that recombination is of the greatest advantage when the double mutant is more advantageous than either single mutant, when the mutant effects are small, when mutations occur with high frequency, and when the population is large.
The Fisher-Muller model presents an argument for how sex can be beneficial through eliminating competition among favorable mutations that have arisen in different genetic backgrounds, instead bringing these together through recombination, speeding up adaptation. Later, Muller shifted focus from favorable mutations to deleterious ones, perhaps inspired by the fact that most non-neutral mutations are in fact deleterious (Loewe and Hill, 2010).
Muller (1964) introduced the concept later dubbed Muller's ratchet, a process in which, in the absence of recombination, deleterious mutations accumulate in a population in an irreversible manner. Without recombination, assuming that back-mutations are rare, Muller argues that offspring will at minimum carry the same mutational load as their progenitors. He proposed this mechanism as a further reason for why sex might win out over asexual reproduction, as sexually reproducing organisms avoid the ratchet due to sex's ability to recreate high-fitness individuals by bringing together unmutated alleles in the same individual. Experiments have shown that ratchet is fastest, and thus the advantage of sex the largest, when mutation rates are high, selection is weak, the organisms (genomes) are large and the size of the population is small (Bell and Graham, 1988).
Kondrashov (1988) complements the work of Muller by introducing the role of synergistic epistasis. This is the idea that most mutations are only slightly deleterious individually but that the cumulative effect of mutations has an increasingly large effect on fitness as the total number of mutations increases (see Figure 2). In a sexual population, some of the individuals born will have many mutations, while some will have few. Since there is a major selective disadvantage to individuals with more mutations, these individuals die out. Sex compartmentalizes the deleterious mutations, and rids the population of them.
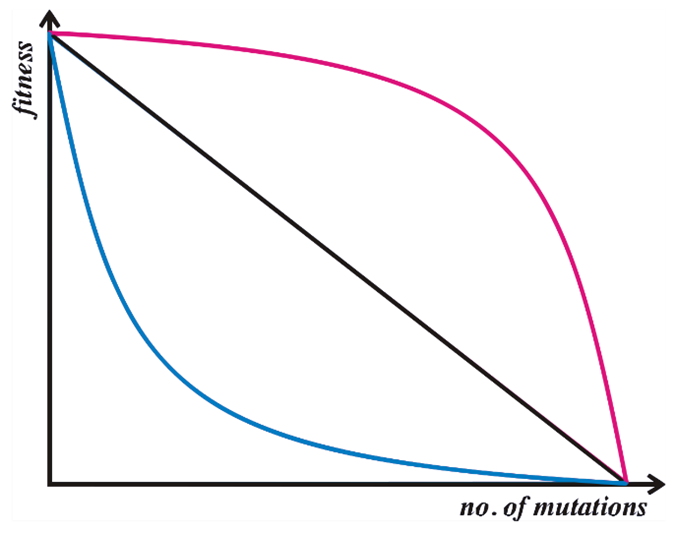
Figure 2: Illustration of different relationships between the number of mutations and fitness. The red line represents synergistic epistasis, where each subsequent mutation has a disproportionately large effect on the organism's fitness. This is a requirement for Kondrashov's model. The other two lines illustrate additive (black) and antagonistic (blue) effects of the number of mutations on fitness (Kondrashov, 1988). Original illustration by MyvReeve, licensed under the Creative Commons Attribution-Share Alike 3.0 Unported license
The ideas presented so far seem to argue that sex persists because it benefits populations by bringing advantageous combinations of genes together, and by eliminating deleterious ones. Mystery solved? Many would argue no. One reason for this is that major parts of the Vicar of Bray thinking can essentially be deemed group selectionist, due to many of the benefits put forth mostly accruing to the group, and not the individual (at least not in any immediate sense). Given that there is a strong consensus that natural selection works on the level of the individual (or even the gene), and not on the species or group level, this poses a problem. Natural selection is not going to promote genes that benefit the species but hurt the individual, because such genes will be outcompeted by non-self-sacrificial variants (Maynard-Smith, 1978). Furthermore, Muller’s ratchet, the Fischer-Muller model, and other forms of reducing selective interference are slow processes. Their benefits do not seem to counter the large costs of sexual reproduction in the short term (Ridley, 1994). Convincing arguments for how sexual individuals can be better off than non-sexual individuals are therefore needed.
2.2 The Ecological Perspective
In the attempt to answer how sex sufficiently benefits the individual despite its two-fold cost, many biologists shifted focus towards ecological explanations. Williams (1975) argued that the genetic mixing inherent to sex, might prove beneficial when the environment is changing rapidly. If the environment changes from generation to generation, the genotype advantageous in one generation might not be advantageous in the next. The much greater diversity of sexually produced offspring compared to asexual ones, could therefore be a good strategy. However, if sex' purpose mainly were as an adaptation to highly variable environments, one would predict sex to be more prevalent in highly fluctuating environments such as high latitudes and altitudes. Generally speaking, this is not what is found. In fact, data suggest sex is actually more common in stable environments such as lakes and tropical rainforests (Bell, 1982). Williams' theory is not without value, but it seems to fall short as a single explanation for the persistence of sex.
Another ecological argument for sex is the Tangled Bank hypothesis. The Tangled Bank holds that since the state of the environment varies spatially even at fairly local levels, different genotypes will be optimal at different locations. Given that each location will only support a limited number of individuals, clonal offspring from an asexual parent will compete intensely with one another for the same set of resources. The offspring of a sexual female, however, by virtue of their greater diversity, will be able to exploit a much wider range of sites as well as different niches in the same location. This might lead to sexual lineages competing less amongst themselves and thus to greater overall success compared to asexual lineages despite its lower reproductive efficiency (Bell, 1982). An elegant idea, but empirical support for the Tangled Bank hypothesis as the driving cause for the persistence of sex is not very strong (Lane, 2010).
Lastly, a promising explanation of how sex can be favored by natural selection is referred to as the Red Queen hypothesis. This ecological hypothesis is named after a character in Lewis Carroll's Through the Looking Glass, who must "do all the running [she can do] in order to stay in the same place". Whilst the Red Queen term is used by biologist as a general reference to perpetual arms races between different species, it is perhaps most strongly associated with the evolution of sex [4]See e.g. Ridley (1994).
The hypothesis states that species are in evolutionary arms races with other species, and particularly parasites rapidly evolving to circumvent its hosts’ defenses. This produces a continually shifting adaptive landscape for the host organism, and in this context, sex can be a winning strategy. Recombination allows for the continual arrival of new combinations of alleles to which the parasite is not well-adapted. In the context of arms races, sexual reproduction and its mixing of genes can prove more advantageous than asexual reproduction and its slower production of novelty. Support for this can be found from the observation of parasite co-evolution in populations of mud snails (Potamopyrgus antipodarum), which have both sexual and asexual genotypes. Studying these populations, it was found that the populations with higher exposure to parasites shifted towards higher frequencies of the sexual genotype (Vergara et al., 2014). These studies, and others, suggest that the evolutionary benefits of the genetic mixing produced by sex can compensate for its costs.
The Red Queen hypothesis provides a strong argument for the primary purpose of sex being that of keeping parasites at bay. There are however doubts as to whether the threat of parasites is a strong enough force to explain the powerful pervasiveness of sex. For example, Howard and Lively (1994) used computer simulations to (among other things) study the role of parasites in explaining the rarity of obligate parthenogenesis in natural populations. They found that their models required that parasite transmission rates were very high, and that parasites had severe fitness effects on their hosts (>80% loss of fitness), in order for sex to gain a decisive advantage over parthenogenesis. Conditions like these are unquestionably present in a number of natural systems, but in no way all.
Om forfatteren
 Forfatter Mads S. Michalsen fullførte sin bachelor i Biologi ved UiB i 2019. Han var fra før av ferdig utdannet sivilingeniør ved NTNU og tok bachelor i Biologi hovedsakelig på grunn av ren og skjær nysgjerrighet og personlig interesse for feltet. Mads er nå bosatt i Oslo, hvor han jobber som management consultant i PwC.
Forfatter Mads S. Michalsen fullførte sin bachelor i Biologi ved UiB i 2019. Han var fra før av ferdig utdannet sivilingeniør ved NTNU og tok bachelor i Biologi hovedsakelig på grunn av ren og skjær nysgjerrighet og personlig interesse for feltet. Mads er nå bosatt i Oslo, hvor han jobber som management consultant i PwC.
På spørsmål om hva som motiverte han til å skrive om akkurat dette temaet, svarer Mads følgende:
Spørsmålet om hvorfor sex (eller nærmere bestemt: kjønnet formering som reproduktiv strategi) i det hele tatt er en greie, er en tematikk jeg ønsket å utforske dypere rett og slett fordi det representerer min kanskje favorittsjanger av problemstillinger: den typen hvor svaret ved første øyekast kan virke ganske så åpenbart, men hvor det ved en nærmere titt gradvis vokser fram et solid mysterium, i dette tilfellet en ordentlig evolusjonær gåte. Det er også artig å gjøre en scientific review av et område som har århundrelang historikk, men hvor det fortsatt pågår høyst levende diskusjoner og langt ifra er satt to streker under svaret. I tillegg er det en tematikk som gir veldig god trening i å holde tungen beint i munnen med tanke på praktisering av robust, evolusjonær logikk.
{kind=link}
1 Response
[…] M. S. (2021). Let’s talk about sex: Exploring evolutionary explanations for the persistence of sexual reproducti…. Bikuben […]